09 非洲起源說:DNA與人類史
克林斯和帕博最擔心的事情是污染。在定序古代DNA上有過許多例子,原先以為成功了,後來卻發現這些古代樣本曾遭到現代來源污染,得出的序列其實是錯誤的。我們每天都會有大量皮膚細胞脫落,許多DNA隨之進入周遭環境,天曉得最後到了哪裡。克林斯想用聚合酶連鎖反應來擴增他想找的線粒體DNA片段,但聚合酶連鎖反應非常敏感,即使只是一個分子都會引發作用,擴增它所遇到的任何DNA,無論這個DNA的來源是古代生物還是現代的活生物。萬一尼安德特人的DNA降解得太厲害,以致聚合酶連鎖反應無法對它發揮作用,卻因為克林斯身上脫落的微小物質污染了古代樣本,導致聚合酶連鎖反應還是發生了,但擴增的卻是污染源的DNA序列,那怎麼辦?如此一來,克林斯或許就得解釋一下他和尼安德特人為什麼剛好有相同的線粒體DNA序列。這對這位年輕人的頂頭上司來說,肯定不是一個令人開心的結果,更不用說他的父母了。為了確保這種事不會發生,克林斯和帕博找了美國賓夕法尼亞州立大學的史東金(Mark Stoneking)實驗室來依樣重作他們的研究。那裡也可能會有污染,但是至少不會是被克林斯的DNA污染,畢竟克林斯遠在另一個大陸上。如果兩個實驗室從這個樣本獲得相同的結果,就可以合理假設他們找到了真正的尼安德特人序列。
關於人類的變異還有一個與我們的直覺極相反的發現:無論這些變異有多少,大多數都跟種族沒有直接的關連。在肯恩和威爾遜證明人類離開非洲的時間出乎意料地晚之前,一般認為,不同的族群已在不同的大陸上互相隔離了無數年代,甚至長達200萬年。依照泡令-祖克坎德爾模型,孤立的族群之間遺傳變異的程度,跟他們隔絕的時間長短有連帶關係,如此一來應該會累積大量的遺傳差異。但是,根據肯恩和威爾遜的結論,我們共同的祖先生存的年代離現代並不遠,所以地理位置相互隔離的族群顯然沒有足夠時間發展出大量變異。因此,儘管膚色等遺傳差異在不同族群之間顯得很明顯,但跟特定種族有關的遺傳差異一般非常有限(即界定族群之變異並不多)。事實上,我們少量的變異大多相當均勻地分佈在不同族群之間:我們在某個非洲族群找到特定遺傳變異的幾率,跟在某個歐洲族群找到相同變異的幾率差不多。這讓人不禁猜測,人類大多數的遺傳變異是在離開非洲之前,就已經在非洲大陸發生,因此拓殖到世界其他地方的族群,才會都有這些變異。
由於這個結果太過驚人,金和威爾遜覺得自己有必要為遺傳進化速率(慢),和身體構造及行為進化速率(快)之間的明顯差異提出解釋。既然遺傳變化這麼少,為什麼動物園裡的黑猩猩和在玻璃窗外觀看它們的人類之間,還會有這麼大的差異?他們的看法是:大多數重要的進化改變,都是發生在控制基因啟閉的DNA片段。如此一來,一個微小的遺傳變化有可能造成重大的影響,例如這個小變化可能改變了基因表現的時機。換句話說,大自然可以藉由調控相同的基因,讓它們以不同的方式運作,從而創造出兩種長相非常不同的生物。
雖然這副在德國出土的骨骸是最早被正式命名為尼安德特人的,但現在已經確認,更早在比利時和直布羅陀發現的骨骸也屬於相同的物種。一個世紀后,更多的尼安德特人骨骸出土。現在的看法是,尼安德特人曾在歐洲、中東各處,以及北非部分地區定居,直到大約3萬年前左右。尼安德特人之所以會有愚蠢笨拙的形象,大多是法國古生物學家布勒(Marcellin Boule)所造成的。他重建尼安德特人的材料取自法國的聖沙拜爾(La Chapelle-aux-Saints),而且是根據單一骨骸作出的,這副骨骸最後被發現是屬於一位患有關節炎的老人的。事實上,尼安德特人的腦稍微比我們大一點(而且因為顱骨較平,所以形狀跟我們不同)。埋葬地點的證據顯示,尼安德特人的文化先進,已經有埋葬儀式,因此他們可能已經有來生的觀念。
住在緯度較高的地區,為什麼會使人失去黑色素?最好的解釋是,這與維生素D3的合成有關,D3的合成過程是在皮膚內進行,而且需要紫外線。D3是鈣質攝取的重要成分,而鈣質則是使骨骼強的重要成分。(D3不足會造佝僂病和骨質疏鬆症。)在我們的祖先離開非洲到季節分明的環境后,全年的紫外線輻射比較少,所以自然選擇有可能對膚色較白的變異有利,因為他們的皮膚內阻擋陽光的色素較少,在可用的紫外線有限的情況下,能以較高的效率合成D3。相同的邏輯也適用於我們在非洲大陸上遷移的祖先,例如南非的桑人,南非的紫外線強度跟地中海差不多,桑人的皮膚卻特別白。似是住在幾乎沒有陽光的北極和附近地區、膚色卻深得令人驚訝的因紐特人,又要怎麼解釋呢?由於氣候因素,他們必須隨時用衣服包住全身,這應該會進一步縮減他們製造維生素的機會。但是,事實上,在他們身上似乎看不到對淺膚色有利的自然選擇壓力,原因可能在於他們已經自行解決D3的問題:他們的飲食中富含魚肉,而魚肉正是這種重要養分的豐富來源。
如同克林斯在10年後對尼安德特人所作的分析,肯恩和威爾遜也是利用線粒體DNA。這麼做的原因有好幾個,但是,就像一般的情況,實際執行上的問題是最重要的考慮。在聚合酶連鎖反應技術成為研究主流之前,要取得足夠的DNA以研究某個基因或區段,是相當讓人頭疼的事。肯恩和威爾遜的研究必須分析147個樣本,而不只是一個而已,因此他們需要用上所有找得到的DNA。人類組織樣本的線粒體DNA比細胞核里的染色體DNA多很多,但是他們仍需要許多組織,才能取出足夠的線粒體DNA。他們的解決方法是使用胎盤。醫院在接生嬰兒后通常會把胎盤丟掉,而它們正是線粒體DNA的豐富來源。肯恩和威爾遜需要做的是說服147位孕婦,把寶寶的胎盤捐出來供科學研究之用。事實上是146位,因為金很樂意捐出她女兒的胎盤。他們知道要使重建的人類族譜儘可能完整,他們所使用的組織就必須取自遺傳多樣性最大的捐贈者群中。在這方面,像大熔爐般的美國族群明顯具有優點:他們不必遠到非洲去找非洲人的DNA——非洲人的基因早已進入美國。但是他們得靠新幾內亞及澳洲的合作夥伴去尋找願意提供組織的原住民婦女(她們在美國的基因庫中所佔的比例並不高)。
兩批獨立的資料,卻對人類的歷史有類似的發現,這是極為驚人的成果。在只研究一個區域,例如線粒體DNA時,所獲得的結果即使很有意義,仍不能當做定論;這個模式所反映的,可能只是此特定區域的DNA歷史所具有的特色,而不是某個重大歷史事件對人類這個物種整體的影響。最重要的是:當「族譜樹」的分支匯聚到一點——所謂的「趨同點」(convergence point),也就是我們研究的所有序列最近代的共同祖先、我們所有人的曾曾曾曾……祖母——這並不必然與人類歷史上任何特定事件有關。雖然它有可能暗示現代人類的起源,或其他一些具有歷史意義的人口學上的事件,但是,以人類歷史的宏觀角度而言,它也有可能只是顯示出一些更為瑣碎的事,例如過去自然選擇對線粒體DNA的影響。然而,如果相同的改變模式可見於基因組中一個以上的區域,就很有可能代表真的找到了過去某個重要事件的遺傳痕迹。
威爾遜先前是用蛋白質來分析人類與猿猴在進化上的分歧時間,等到DNA技術成熟時,他將進一步的分析工作交給他指導的研究生瑪麗—克萊爾·金(Mary-Claire King)。她在1975年寫出的報告,是20世紀最傑出的科學論文之一。其實有很長一段時間,這個研究似乎不太可能有很好的結果,特別是從金的觀點而言。她的研究進行得並不順利,部分原因在於70年代初期伯克萊的反越戰運動讓人很難專心。金一度考慮到華府去為消費者保護健將奈德(Ralph Nader)工作,但幸好她聽了威爾遜明智的建議:「如果每個做實驗失敗的人都不再搞科學,世上就不會有科學存在。」金決定繼續她的研究。
最近一個針對哥倫比亞人Y染色體與線粒體DNA變異的研究,證明了類似的效應。在大多數的社會族群中,哥倫比亞人的Y染色體和西班牙人的Y染色體相同,這是征服南美洲北部沿岸西班牙屬地的歐洲人留下來的生物遺產。事實上,在接受檢測的Y染色體中,大約94%都是源自歐洲。似有趣的是,線粒體模式卻相當不同:現代哥倫比亞人擁有多種類型的美洲原住民線粒體DNA。這樣的結果所代表的含義相當清楚:入侵的西班牙人(當然都是男性)娶了當地的女人。哥倫比亞人沒有美洲原住民的Y染色體類型,這透露出殖民時代的滅族大屠殺悲劇:當地的男性被消滅,而女性則被征服者通過性的方式「同化」了。然而,有時這種XY來源長久以來不對稱的情況是因為文化延續的問題,而不是不同文化激烈衝突的結果。印度的少數民族帕西人(Parsee)認為自己是祆教徒(Zoroastrian)的後裔,祆教徒屬於印歐民族雅利安人,在7世紀時因宗教迫害逃離波斯(今伊朗)。對現代帕西人所作的遺傳分析顯示,他們真的保有伊朗人的Y染色體,但他們的線粒體DNA通常是印度型的。在這個例子中,這種不對稱性是因為傳統而維持了下來:只有信奉祆教的帕西人所生的兒子,才會被視為真正信奉祆教的帕西人。因此帕西人社群里的成員都經由父系繼承到Y染色體。遺傳學確認了傳統的影響力。

古代與現代的烤肉聚會:上圖是畫家筆下3.5萬年前歐洲南部一個尼安德特人的營地,下圖則為現代的類似場景(20世紀50年代的美國家庭)。我們是尼安德特人的後代嗎?DNA的證據顯示不是。
但是我們要怎麼從這個「分子鍾」看出「時間」呢?換句話說,我們要怎麼測量我們和自然界其餘生物的遺傳差異?在60年代晚期,早在定序DNA的時代來臨前,加州大學伯克萊分校古怪的紐西蘭人威爾遜(Allan Wilson)和他的同事薩瑞奇(Vince Sarich)就已開始用泡令和祖克坎德爾的邏輯來研究人類和人類的近親。當時,定序蛋白質仍然相當棘手read.99csw.com費力,但是威爾遜和薩瑞奇卻找到一個巧妙的快捷方式。
這個模糊不清的問題一直糾纏著肯恩和威爾遜的研究結果:這究竟是進化的修補過程所造成的,還是在人類史前時代整體發展中的某一個重大事件所造成的?但是當安德席爾也在Y染色體上觀察到類似的模式時,這個模糊性便消失了。兩者的一致,強烈顯示在15萬年前,人類族群的確經歷了一場劇烈的遺傳變化,而且其劇烈程度已經足以同時對線粒體DNA和Y染色體造成影響。這當中涉及的現象即稱為遺傳瓶頸(genetic bottleneck),我們稍後將進一步討論這點。
我們的線粒體DNA都是從母親繼承而來。父親的遺傳物質包在精|子的頭部,其中並不包括線粒體的物質。精|子的DNA被注入已含有母方線粒體的卵細胞內。因此,肯恩和威爾遜追蹤的是人類母系的歷史。由於線粒體僅在母系中傳承,所以沒有機會重組。所謂重組也就是染色體的臂部片段互相進行交換,所以突變可以從一個染色體移動到另一個染色體上去。在根據DNA序列的相似性重建族譜時,線粒體DNA沒有重組的情形成為一大優點。如果兩個序列有相同的突變,我們就知道它們一定有共同的祖先(也就是最早產生這個突變的生物)。如果有重組發生,這些譜系的突變就有可能是從剛剛發生的重組活動中獲得的,所以擁有相同的突變並不一定代表有共同的祖先。因此,利用線粒體DNA來建立族譜的邏輯很簡單:類似的序列(有許多相同突變的序列)表示親緣關係近;有許多差異的序列表示親緣關係比較遠。從族譜的樹狀示意圖(P185)來看,親緣關係近的物種(由年代較近的共同祖先傳承下來的物種)在「族譜樹」上的距離較近;親緣關係較遠的物種分散得比較遠,因為它們的共同祖先存在的年代比較久遠。
在生化層面,我們也不是很清楚人類有哪些相對的優點,目前為止,我們只知道人類和黑猩猩之間有兩個差異。第一個差異是:兩個物種的細胞外圍都具備的一種糖分子,即唾液酸(sialic acid),在黑猩猩體內,這個分子通過一種酶的作用,會有些微變化;但是在人類體內,為這個酶編碼的基因總是發生突變,亦即沒有酶生成,而人類細胞表面的唾液酸也就沒有變化。這個差異是否具有任何重要意義,我們不得而知。第二個差異在於FOXP2基因。這個在2002年由帕博的小組所發現的差異,可能更有意義。目前已知FOXP2基因與人類的語言有關。(由於已經發現FOXP2基因發生突變會引起語言障礙,因此這個基因被媒體稱為「文法基因」[the grammar gene],但這個詞容易引起誤解。)在FOXP2蛋白質有715個氨基酸的鏈上,人類只有兩個氨基酸和黑猩猩及大猩猩不同,黑猩猩和大猩猩的FOXP2蛋白質則是完全相同的。事實上,這些氨基酸在所有試驗過的哺乳動物中都是一樣的,只有人類不同。此外,對這個基因內部及周圍的DNA變異模式所進行的統計分析顯示,在人類進化期間,自然選擇可能對此蛋白質進行了某種塑造。這讓人不禁產生一個(尚且言之過早的)想法:FOXP2可能是進化上的確鑿證據,就像槍擊案中還在冒煙的槍,讓人得以窺知語言起源上的關鍵步驟。
偉大的化學家泡令是從分子層面來研究進化的先驅。在20世紀60年代早期,他和分子進化學家祖克坎德爾(Emile Zuckerkandl)比較了數個物種之間相應蛋白質的氨基酸序列。當時正值蛋白質定序的初期階段,數據自然相當有限。但是他們注意到一個驚人的模式:兩個物種在進化上的親緣關係愈近,他們相應的蛋白質序列愈相似。以血紅素分子的一個蛋白質鏈為例,他們發現,在它全部141個氨基酸中,人類和黑猩猩之間只有一個氨基酸不同,但人類和馬卻有18個氨基酸不同。分子序列的數據反映出馬和人類在進化上的分歧出現得比人和黑猩猩要早。找出隱藏在生物分子里的進化歷史,現在是常見的做法,但在當時這卻是具有爭議的新概念。
這個尼安德特人序列跟現代人線粒體DNA序列之間的共同點,比它與黑猩猩之間多,這說明尼安德特人無疑是人類進化譜系中的一支。但在此同時,尼安德特人的序列,和克林斯拿來作比較的全部986個現代人線粒體DNA序列之間,也有驚人的差異。在這當中,即使是跟尼安德特人的序列最接近的現代人線粒體DNA序列,至少也有20個鹼基對(佔5%)是不同的。後來,另外兩個尼安德特人(分別在俄羅斯西南部和克羅埃西亞出土)的線粒體DNA也被定序,而且如同預期,這兩個序列跟首先被定序的那一個並不完全一樣(尼安德特人之間自然也會有變異,如同現代人之間),但它們很類似。
漢默和其他人利用Y染色體來追蹤猶太人的大流散過程,獲得許多有趣的結果。例如在歐洲定居了1200年(現在也居住在美國和其他地方)的徳系猶太人(Ashkenazim),至今仍具有源自中東的遺傳特徵。事實上,分子研究已清楚顯示,至少在遺傳上,猶太人和所有其他的中東族群(包括巴勒斯坦人)毫無區別,經書上也是這樣記載的。據說偉大的希伯來人族長亞伯拉罕(Abraham)跟不同的妻子生育了兩名兒子:以撒(Isaac)和以實瑪利(Ishmael)他們兩人分別是猶太人和阿拉伯人的祖先。同一人的後代競然成為不共戴天的死敵,這在基因證實了傳統說法后,更顯諷刺。
黑猩猩是親緣關係跟我們最近的動物,它們濃密毛髮下的皮膚大多都沒有色素——黑猩猩可以說是擁有一身白皮膚。在500萬年前,人類和黑猩猩的共同祖先可能也同樣是白皮膚,因此我們可以推論,現在非洲人(以及出生在非洲的最早的智人)特有的深膚色是在後來人類的進化過程中出現的。在失去毛髮后,色素成為保護皮膚細胞不受陽光中紫外線輻射傷害的要素。
卡瓦利-斯弗扎知道,只有由父傳子的基因,亦即人類基因組中由父系傳承下來的一些成分,才是肯恩和威爾遜對人類進化看法的最有力證明。如果能藉由追蹤父系,亦即不是走肯恩和威爾遜分析線粒體DNA的母系路徑,而是走父系路徑來獲得相同的結論,就可以作為真正獨立的確證。在基因組中,男性特有的成分自然就是Y染色體。根據定義,擁有Y染色體的人即是男性(要記得,Y染色體是男性自父方繼承而來的,男性的精細胞不是含有X染色體,就是含有Y染色體;精|子會跟永遠只含有X染色體的卵細胞結合,因此精|子便決定了我們的性別,XX組合會產生女性,而XY的組合是男性)。Y染色體握有了解男性遺傳歷史的關鍵。此外,由於重組只發生在成對的染色體之間,使用Y染色體不會碰上重組的問題,可以避免進化分析上的可怕陷阱。任何Y染色體都是獨一無二的,絕不會有另一個Y染色體可以跟它交換遺傳物質。
有一次到愛爾蘭時,我特別到蒂珀雷里郡(Tipperary)的檔案室,尋找更多與我外曾祖父有關的資料。那個檔案室位於尼納(Neneagh),離利默里克郡(Limerick)大約20英里,先前是一所監獄。但我的偵探工作一無所獲,那裡完全找不到有關格雷鎮的記錄,我只能判斷說,在我那位可能不識字的祖先的墓碑上所刻的市名是想像出來的。因此,我結束了惟一一次針對自己家譜的研究。一直到最近,我對族譜的興趣才又被勾起來。現在人類族譜的架構已經由肯恩和其他人建立起來,我很想知道自己在人類族譜里的位置。塞克斯的「牛津尋祖」這類公司,代表了家譜研究的新領域。他們用高科技的實驗室取代布滿灰塵的檔案。牛津尋祖公司在取得我的DNA樣本后,開始進行線粒體DNA和Y染色體的分析。只可惜檢驗結果顯示,有關我祖先的歷史一點也不浪漫,他們也不是來自遙遠的異國。我真的跟我所害怕的一樣,主要是蘇格蘭人與愛爾蘭人的產物。我甚至不能把我比較野蠻的一面,歸咎於古代時曾有維京人侵入我的家族血統。
從遺傳證據來看,桑人似乎在最初遷移至非洲南部和東部以後,在整個歷史中都處於相對孤立的狀態。從桑人罕有的(至少在我聽來很稀罕)「喀嗒」(click)語言來看,社會語言學上的證據也足以支持這種推論。由於說班圖語(Bantu)的族群在1500年前開始從中非西部向外擴張,所以桑人目前的分佈範圍極為有限。班圖人的擴張迫使桑人遷至貧瘠的邊陲地區,例如卡拉哈里沙漠(Kalahari Desert)。
若說有哪種性狀在人類族群中的分佈情況是很難了解的,那就是乳糖不耐症(lactose intolerance)。哺乳動物的乳汁,包括人類的,都富含一種糖類,稱為乳糖;而初生的哺乳動物一般會製造一種特殊的酶,即乳糖酶(lactase),它會分解腸道中的乳糖。但是在斷奶時,大多數哺乳動物,包括人類(至少包括大多數非洲人、美洲原住民和亞洲人),會停止製造乳糖酶,因此成人無法消化乳糖。有乳糖不耐症的人喝一杯牛奶就會引起腹瀉、腸氣和腹脹等不適癥狀。但相對地,大多數高加索人和其他一些族群的人卻終生都會製造乳糖酶,因此終生都可以食用乳製品。

「線粒體夏娃」成了封|面|女|郎:1988年1月11日出刊的美國《新聞周刊》以「尋找亞當和夏娃」的標題,報道威爾遜等科學家對人類起源的爭議性理論。配合「非洲起源說」,亞當和夏娃以非洲人的面貌現身說法。
從桑人相對比較穩定的歷史來看,他們是否可以讓我們窺知所有現代人的祖先是什麼模樣?有可能,但不必然。在過去的15萬年間,桑人的譜系也可能發生過重大改變,與祖先不同了。即使想從桑人來推測我們早期祖先的生活方式,也是有疑問的:自從說班圖語的族群在較晚的年代遷移到這裏后,桑人退居沙漠一隅,而他們現在的生活方式是適應這種嚴酷環境的結果。2000年,我拜訪卡拉哈里沙漠的一個桑人小區,體驗了幾天他們獨特刺|激的生活。我對他們驚人的實用主義生活方式感到相當震撼,他們以有效率又實際的方式執行眼前的所有工作,即使那些不是他們平常會做的事也一樣,例如修理漏氣的輪胎。那時我真希望我有更https://read.99csw.com多適應能力可以跟他們一樣好的同事!從遺傳觀點來看,縱使他們跟我的遺傳相差十萬八千里,但我們竟然在心性上這麼相似,實在令我震驚。
桑人獨特的遺傳與文化特色不久就會消失。卡拉哈里沙漠的年輕人不怎麼想跟他們遊牧的父母一樣,過著簡單的採集/狩獵生活。例如我去拜訪桑人族群時,他們表演了一場傳統的「神靈附體之舞」(trance dance),但是年輕的族人顯然對長老們古怪的舞姿感到困窘。他們遲早會搬離自己的小區,跟其他族群通婚。
遺傳分析也讓我們得以重建比較後期的史前歷史。例如漢默已經證明現代的日本人,是古代以採集/漁獵為生的繩紋族(Jomon)和相對比較近代的彌生族(Yayoi)混種后的結果。繩紋族目前的代表族群是蝦夷族(Ainu)原住民;彌生族大約在2500年前從朝鮮半島抵達日本,帶來了編織、金屬工藝,以及以稻米為主的農業。我們也在歐洲看到一波波遷移的證據,這些遷移大多與農業技術的進步有關。巴斯克族(Basques)和凱爾特族(Celts)在遺傳上與歐洲其餘的族群不同。有一個解釋是,這些族群面對更晚抵達的族群時,被迫遷移至相對較偏遠的地區。(巴斯克人現在居住於法國和西班牙邊境的比利牛斯山脈山區;凱爾特人較晚抵達歐洲,分佈於歐洲西北緣,從法國的布列塔尼[Brittany]一直到愛爾蘭和不列顛西部地區。)
從免疫系統對外來蛋白質的反應強度,可以看出那個外來蛋白質跟被入侵的生物體之間有多少差異:如果它和這個生物體本身的蛋白質相當類似,則免疫反應也會相對較弱;但是,如果這兩個蛋白質差異很大,免疫反應也會按比例變得較強。威爾遜和薩瑞奇從一個物種取出一種蛋白質,再測量它在其他物種體內引起的免疫反應,以此來比較反應強度。他們藉此建立兩個物種之間分子差異的指標,但是要讓這個「分子鍾」具有時間意義,他們必須實際測量。
令大家驚訝的是,這個爭議已經由DNA解決:1997年從1856年出土的骨骸中取出的DNA,已有3萬年之久。DNA的進化正是為了安全地保存信息,使其能代代相傳,因此它的化學穩定性高,不會自動降解,也不會輕易和其他分子起作用。但它並非不會受到化學破壞。在死亡時,身體里的遺傳數據會跟其他的成分一樣,變得容易受許多降解者影響,這些降解者包括化學反應物質,以及能分解分子結構的酶。這些化學反應需要水,因此如果屍體脫水的速度夠快,DNA就有可能保存下來。但是即使在理想的保存狀況下,DNA分子可能頂多隻能保存5萬年。因此,要從已經有5萬年之久、保存得並不好的尼安德特人骨骸里取出易讀的DNA序列,實在是很不容易的事。
亞利桑那大學的漢默(Michael Hammer)和這個領域的許多大師(包括肯恩和帕博)一樣,也是在威爾遜的伯克萊實驗室接受訓練。雖然漢默最初感興趣的是老鼠,但肯恩和威爾遜發表的線粒體DNA研究,讓他從嚙齒動物轉向研究人類的歷史。Y染色體的信息對肯恩和威爾遜的整個假說是很重要的試驗,他是最早了解到這一點的人員之一。但是起初Y染色體不願輕易讓人窺知它的秘密。由吉爾伯特實驗室所作的一項研究,把取自許多人Y染色體的同一個DNA區段定序,結果只發現這些區段的序列一模一樣,辛苦研究半天,卻沒獲得任何有關遺傳上相互關係的信息。然而漢默堅持研究下去,最後他和其他人一起把Y染色體變成人類學的金礦,他們的成就在安德席爾發表那篇重要論文時達到了顛峰。
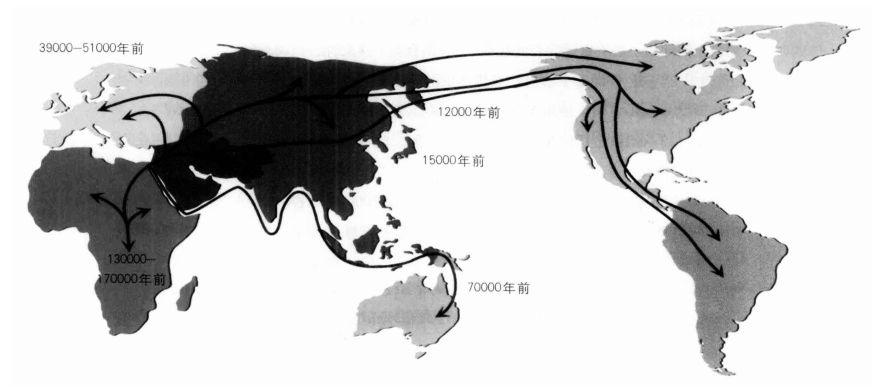
遠離非洲:人類起源於非洲,並從非洲散播至世界各地。圖中的移民年代是根據線粒體DNA的資料推估出來的結果。
傳統也與猶太人的遺傳變異模式有關。最近一項研究已經證明,古代猶太人的祭司階級kohanim及其後代(至今仍可由其姓氏柯恩[Cohen]來辨別)所擁有的Y染色體相當獨特,足以使他們自成一個族群。即使居住地區最偏遠的猶太族群,也就是在古代猶太人被逐出故土的「大流散」(Diaspora)中遷移至最遠方的猶太族群,例如南非的蘭巴(Lemba)。柯恩型Y染色體仍保存下來,幾乎就像神聖的宗教經文一樣。一般認為這種染色體的來源始於亞倫(Aaron);根據聖經記載,亞倫是祭司階級的創始人,也是摩西的兄長。柯恩Y染色體序列真的源自亞倫,並在父子代代相傳下保持完整,也並非不可能的事。在猶太人的歷史中,這樣的傳統一直被嚴格保存下來。
現在回頭來看我們虛擬族群中Brown, Browne, Bowne, Frown或Broun這幾個姓氏。我們可以推論,他們全都源自起始三個譜系中的同一個譜系:Brown。那麼人類線粒體DNA和Y染色體的數據所具有的含義,應該也不會讓我們驚訝:15萬年前,有許多不同的線粒體DNA序列和許多不同的Y染色體序列,但是今天的序列全都是其中一個線粒體DNA序列和其中一個Y染色體序列的後代。所有其他的序列都已經滅絕,大多數可能是在某個古代的「瓶頸事件」中消失的,這事件有可能是族群人口因瘟疫而銳減、氣候改變或任何其他原因。但是無論我們早期歷史中的這個災難事件是什麼,有一件事很清楚:在一段時間后,我們的祖先族群開始離開非洲,展開人類殖民全球的漫長旅程。
人口統計學上的因素對族譜有什麼影響?任何系譜都是不同的血統相互消長的結果:隨著時間流逝,有些會繁衍,有些會滅絕。以姓氏為例,假設在1000年前,某個偏遠小島上原本只有三種姓氏:Smith, Brown和Watson。再假設,當要把新生兒的名字抄到出生記錄簿上時,偶爾會犯一些小錯誤(即突變)。這些錯誤很罕見,也很輕微,所以我們還能夠分辨這個被寫錯的姓氏原本是哪一個:Browne顯然是Brown的突變。現在假設過了1000年,到了現代,我們發現島上的姓不是Brown,就是Browne, Bowne, Frown或Broun. Smith和Watson這兩支已經滅絕,而Brown這一支成功繁衍下來(而且通過突變而變得多樣化)。這代表發生了什麼事?完全是運氣,導致Smith和Watson這兩支滅絕。例如,可能有一代的好幾對Smith夫婦生的大多是女兒。假設島上的姓氏是沿父系繼承的(這是根據傳統所作的假設,儘管現代已有其他的常規),生許多女兒有可能造成下一代中姓Smith的人減少。現在假設下一代的Smith也生育了太多女兒,人口效應就會再度增強,最後會完全沒有了姓Smith的人。Watson這一支的情形也一樣。
由於人類的進化可說歷時甚短,我們在族群中所看到的具有一致性的差異,大多數都可能是自然選擇的產物,例如膚色。
卡瓦利-斯弗扎的同事安德席爾(Peter Underhill)在2000年發表了一篇論文,造成轟動。安德席爾研究的是Y染色體,就像肯恩和威爾遜研究線粒體DNA—樣,他的研究發現跟他們的極其類似。他再度發現人類族譜的根源在非洲,而且也再度證明了人類族譜很短:它不像人類學家原本所設想的,是一棵遠古的高大樹木,而是跟肯恩和威爾遜的分析結果一樣,是一棵灌木,樹齡只有15萬年。
事實上,桑人和其他族群逐漸混合的趨勢已見諸歷史。例如南非共和國黑人領袖曼德拉所屬的科薩部落(Xhosa)就混合了班圖人與桑人,科薩語雖然是以班圖語為主幹,似也混入了許多桑人典型的喀嗒聲。在今日科技突飛猛進的時代,桑人在遺傳與文化上的完整性不太可能再存續多久。幸好過去數十年來有不少研究,其目的都在於了解和記錄這個獨特的民族和他們的生活方式。約翰內斯堡威特沃特斯蘭德大學(University of Witwatersrand)的托比亞(Philip Tobias)不僅是這些研究計劃的發起人,多年來,在種族隔離的黑暗時期,他也擔任桑人的非正式發言人,捍衛他們的權益。口懸河的威爾士人傑金斯(Trefor Jenkins)先是在尚比亞的數個銅礦城鎮行醫,後來搬到南非,他也是長期在遺傳學方面研究桑人和其他原住民族群的先鋒。
Y染色體的重要信息讓我們得以重建人類拓殖新世界的歷史,在整個人類史上,拓殖新世界是相對比較後期的發展階段。目前有關人類最早在美洲的定居地點的說法仍莫衷一是,傳統上獲得最多支持的地點是新墨西哥州的克洛維斯(Clovis),時間約在1.12萬年前;但是支持智利蒙特維爾第山(Monte Verde)的人則聲稱,那裡的遺址至少已經有1.25萬年之久。第一批美洲印第安人是在最後一次冰河時期穿過了白令海峽的陸橋,還是乘船走得比較偏南的路線,也仍有爭議。然而,遺傳數據明確顯示,最早的創始族群很小:由於只找到兩種主要的Y染色體序列,因此當時似乎只有兩個明確的群體,而且每個群體可能頂多隻有一個家庭。在印第安人之間,線粒體DNA變異比Y染色體變異廣泛得多,顯示每個創始群體中女性多於男性。在這兩種Y染色體序列中,比較常見的那一種可能代表第一個創始群體;在第二個群體(包括今日納瓦霍[Navajo]和阿帕切[Apache]的祖先)抵達前,第一個群體已建立了後代族群。比較常見的序列也具有另一個特色:它有一個在世上其他地方極少發現的突變。這個突變是2002年首度發現的,它進一步證明了其擁有者是更早的拓荒先驅,據估計,它已有1.5萬年之久,略早於已知最古老的先民遺址。
我以前對族譜學絲毫不感興趣。但我的家族(我猜許多家族都一樣)有一位專門的檔案保管人,那就是我的貝蒂阿姨。她把一生的時間都花在了解家族成員之間的關係,以及這些關係的來龍去脈上。她發現我們沃森家(源自蘇格蘭低地家族)是1795年首度抵達美國,在新澤西州的卡姆登市(Camden)落戶。她也堅信,林肯在伊利諾伊州春田市的宅第是我父系某位祖先所設計的。但我向來對我的愛爾蘭血統比較感興趣,也就是我外婆的家族。我的外曾祖父母九-九-藏-書在19世紀40年代馬鈴薯大飢荒期間逃離愛爾蘭,最後來到印第安納州。我的外曾祖父葛利森(Michael Gleason)於1899年過世,同年我母親誕生。在我外曾祖父的墓石上刻著,他來自愛蘭爾的格雷(Glay)鎮。
在我撰寫本書時,黑猩猩基因組計劃已經接近完成,等這個計劃完成後,將會揭開金和威爾遜所找到的、構成那1%差異的DNA之秘密。我的猜想是,最終會證明他們的想法是對的:造成關鍵性差異的,不在於基因本身,而在於基因的調控。我懷疑,人類只不過是擁有幾個獨特(而且具有特定功能)的基因開關的大猿罷了。
了解人類史的另一個關鍵,可能在於卡瓦利-斯弗扎等人發揮得淋漓盡致的一項觀察結果:遺傳進化模式經常與語言進化模式有相互關係。當然,基因與語言之間明顯具有一些可以模擬之處:它們都是代代相傳,都會歷經變化。但語言的變化可能特別快,青少年的父母應該都體會得到這一點。同樣地,美式英語和英式英語相同,但是又具有明顯不同之處,儘管它們才各自進化了幾百年而已。因此,我們可以按照各個語言的異同點來重建語言族譜,這跟重建遺傳族譜的方式大同小異。但是在許多實例中,如同達爾文本人所預測的,更重要的一點在於我們可以找出這兩個族譜之間有用的對應關係,如此一來,我們對一個族譜的了解可以加深我們對另一個族譜的了解。凱爾特族和巴斯克族就是絕佳的實例:這兩個族群在遺傳上與歐洲的其餘族群不同,其各自的語言也與歐陸其餘的語言不同。至於在新世界方面,一個具有爭議性的語言學理論指出,美洲只有三個本土的主要語族,其中兩個與用印第安人的Y染色體資料辨識出來的早期遷徙事件有關;第三個語族是三者中最小的語族,則與孤立的因紐特人(Inuit,即愛斯基摩人)有關。
金和威爾遜在比較黑猩猩和人類的基因組時,混合採取了數種方法,包括一種稱為DNA雜合(DNA hybridization,亦稱DNA配對法)的聰明方法。當DNA互補的兩股合併形成雙螺旋時,可以藉由將它們加熱至攝氏95度,把它們分離(此現象在分子遺傳學上稱為「融解」[melting])。如果這兩股並非完全互補,也就是當其中一股有突變時,會是什麼情形?結果發現這兩股會在攝氏95度以下的溫度就融解。至於這個溫度有多低,則取決干這兩股之間的差異程度:差異愈大,要使它們分離所需的熱能就愈少。金和威爾遜用這個原則來比較人類和黑猩猩的DNA。兩者的序列愈相似,其雙螺旋的融點就會愈接近完全互補時的標準融點(攝氏95度)。相似程度的實驗結果令人相當驚訝:金推論出人類和黑猩猩的DNA序列的差異程度僅在1%而已。事實上,人類和黑猩猩的相同程度,比黑猩猩和大猩猩之間還高。黑猩猩和大惺猩的差異程度大約是3%。
個別的考古資料也支持這樣的假設。顯然大約在5萬年前,現代人突然在「文化上」觀代化了:我們從這個時期的遺物中發現,他們首度有了純粹用於裝飾的飾品,開始常規性地使用骨頭、象牙和貝殼來製作日常的實用工藝品,並且也開始改良和採集的技術。當時究竟發生了什麼事?我們可能永遠無法知道。但我們忍不住猜想,或許是語言的發明促成了這一切,以及人類至今所有的成就。
肯恩、威爾遜以及他們的研究夥伴,從此徹底改變了我們對人類歷史的看法。
1856年8月,德國採石場工人在杜塞爾多夫(Du-sseldorf)外的尼安德山谷(Neander Valley)炸開一個石灰岩洞后,發現了一副不完整的骨骸。這些骨骸乍看之下像是屬於一種已絕種的熊,這種熊的骨骸經常出現在洞穴里。但是當地一位教師發現,這副骨骸跟人類的親緣關係近得多。這些骨頭的主人身份成為爭議的焦點,其中尤以頭骨上厚厚的眉脊最令人費解。最奇特的說法是,這些骨骸屬於一位受傷的哥薩克騎兵,他在拿破崙戰爭期間爬進這個洞穴后死亡。這個怪異的理論表明,這個可憐的傢伙原先就有慢性病,老是皺著眉頭,致使額部中間永久凹陷,且讓眉脊特別突出。達爾文的《物種起源》出版后,引起各界對人類起源的熱烈爭論,4年後,也就是1863年時,這些在德國出土的骨骸主人被命名為尼安德特人(Homo neanderthalensis)。他們屬於一種跟智人(Homo sapiens)不同但相似的物種。
但是在德國慕尼黑大學,身材高大、說話簡潔的瑞典人帕博(Svante Pǎǎbo)決定嘗試解決這個問題。如果世上有人能做到,恐怕非他莫屬。帕博是萃取所謂的「古代DNA」的先驅,先前他曾成功地取出埃及木乃伊和冰凍長毛象的DNA;1991年,有5000年歷史的「冰人」因阿爾卑斯山冰河溶化而出土后,他也曾取過冰人的DNA。然而,儘管有這些了不起的成就,要鑽到珍貴的尼安德特人遺骨里尋找完整的DNA,就算真的找得到,也是一項非常艱巨的任務。他的同事,考古學家史密茲(Ralf Schmitz)回憶道:「那就像是拿到割開《蒙娜麗莎的微笑》的許可證一樣。」
接下來,威爾遜的伯克萊實驗室又在1987年拋出一個最具震撼力的炸彈。他和同事肯恩(Rebecca Cann)利用DNA序列變異的模式,建立了整個人類的族譜。這是極少數能登上美國《新聞周刊》封面的科學成果之一。
事實上,這個模式雖然與我們的直覺印象相反,卻很容易解釋。幾乎世界各地所有的傳統社會都有人類學家所謂的父權地域制(patrilocality):來自不同村莊的人結婚後,女性會搬到男性所在的村莊,但男性不會搬到女性原本居住的村莊。假設現在有一個來自A村莊的婦女嫁給B村莊的人,然後她便搬到B村莊去,生了一個女兒和一個兒子。後來,這個女兒嫁到C村莊,這個兒子則把D村莊的婦女娶過來。由此看來,男系一直待在B村莊,但女系在兩代之間從A移到B,再移到C。這個過程代代持續,結果女性的遷移範圍很廣,但男性的遷移範圍則很小。男性的確偶爾會遠離家鄉去征服遠方的土地,但是在人類浩大的遷移模式中,這些事件並不重要:至少在遺傳的層面,人類歷史其實是取決於女性在一個個村莊之間一步步的遷移。
遺憾的是,即使最先進的遺傳學技術,目前也仍無法理清人類文化的起源。考古證據顯示,在進化的第一個階段,我們的祖先的日常活動和其他的類人差不多,包拈尼安德特人在內。以色列的斯虎爾(Skhul)有一個洞穴遺址,其中的證據顯示,大約在10萬年前,現代智人和尼安德特人的族群曾經共同存在,沒有任何一方對另外一方造成明顯的威脅。但是,如同先前所見,現代人在3萬年前左右消滅了眉脊突出的表親尼安德特人。由此看來,似乎在這7萬年期間,現代人因為技術以及(或者)文化上的進步而取得了優勢。

體型對氣候的進化適應。肯亞已經適應炎熱氣候的馬薩伊人(左),以及格陵蘭已經適應寒冷氣候的因紐特人(即愛斯基摩人)。
若說人種之間相對較少的差異很有趣,那麼人類整體的共同特色就更有趣了:我們和親緣關係最近的生物為什麼會有這麼大的差異呢?我們先前已經看到,人類大約在500萬年前和黑猩猩分道揚鑣,但是這段時間僅僅只夠讓我們進化出1%的遺傳差異。不過就在這1%的差異中,有一些關鍵性的突變讓我們成為如今這種會思考、會說話,了不起的生物。其他物種是否具有某個程度的思想能力,尚待討論,但是其他物種都沒有產生達芬奇和克里克這類人物,卻是不爭的事實。
牛津大學的塞克斯(Bryan Sykes)作了許多研究,揭露出現代歐洲複雜的遺傳地圖。傳統看法向來認為,現代的歐洲人大多源自在地中海與波斯灣之間「肥沃的新月地帶」(Fertile Crescent)、發明了農業的中東族群。但是塞克斯發現,歐洲人大多數的祖先不是源自於新月地帶,而是源自於中東人入侵前歐洲本地更古老的族群,以及從歐亞中部(Central Eurasia)遷移過來的族群。這類族群包括分別在公元前500年及公元400年左右,從東方橫掃歐洲的凱爾特人及匈奴人。塞克斯也在進一步闡述他對線粒體DNA的分析結果時,主張所有歐洲人都起源於7個「夏娃之女」(daughters of Eve)之一。所謂的「夏娃之女」是他對歐洲線粒體DNA族譜上少得驚人的幾個主要祖先的稱呼。他還成立了牛津尋祖公司(Oxford Ancestors),你可以付一筆錢,請該公司幫你定序你一部分的線粒體DNA,查清楚你究竟系出這「七仙女」中的哪一女。

亞伯拉罕的兩個妻子各有一子,據說猶太人和阿拉伯人便是這對同父異母兄弟的後代。
根據我們對分子的了解,現在我們已經知道紫外線會引起皮膚癌:它們會讓雙螺旋的相鄰胸腺嘧啶鹼基黏在一起,使DNA分子就像打結一樣。當DNA進行複製時,這個結經常會造成鹼基插錯,從而造成突變。如果在偶然的情況下,那個突變剛好發生在調節細胞生長模式的基因上,就可能造成癌症。皮膚細胞產生的黑色素(melanin)能減少紫外線的傷害。跟我一樣白皮膚的人都很清楚,雖然晒傷通常並不會致命,其對健康的威脅卻比癌症大得多。因此也不難想像自然選擇會有利於膚色變黑,這樣不僅能防止癌症,也能預防嚴重晒傷可能引起的感染。
到紐約市中心曼哈頓的街道上逛一圈,看到形形色|色的人,你會覺得我們人類是地球上遺傳變異最多的物種。但是,以我們已取得遺傳信息的物種而言,相較於其中的大多數,其實人類基因組的變異明顯少得多。就個體之間的差異而言,每1000個人
九*九*藏*書類鹼基對中大約只有1個是不同的。因此在遺傳上,我們有99.9%相同,以其他物種的標準來看,這種差異算是很小的。儘管果蠅看起來都一樣,但它們的變異程度卻比我們高10倍。即使是南極洲上每一隻都長得一模一樣的阿德利企鵝(Adélie penguin),遺傳變異程度也是我們的2倍以上。就連跟我們關係更近的黑猩猩,遺傳變異程度也沒我們這麼少:黑猩猩的遺傳變異程度大約是我們的3倍,大猩猩是2倍,紅毛猩猩則是3.5倍。
基於這些遺傳證據,我們可以下結論說:尼安德特人在人類及其親戚的進化樹上,的確佔有一席之地,但他們那條分支跟現代人這條分支相距很遠。如果3萬年前他們在歐洲相遇時,尼安德特人和現代人真的曾經混種,尼安德特人的線粒體DNA應該會進入現代人的基因庫。但是我們並沒有看到這類輸入的證據,這顯示現代人消滅了尼安德特人,而不是跟他們混種。至於他們是在直接的衝突中消滅了尼安德特人,還是經由比較細膩的方式,就不是DNA可以告訴我們的了。
就算我們對人類的遺傳多樣性還有一丁點自豪,也會在得知下面的事實后消散:人類基因組計劃的結果顯示,我們的DNA中大約只有2%為基因編碼,這意味我們的變異至少有98%是發生在沒有作用的基因組區域。由於自然選擇非常有效率地消滅了對基因組中重要功能部分(例如基因)造成影響的突變,因此突變通常會在非編碼的「垃圾」區域累積。我們之間的遺傳差異很少,而這所造成的不同甚至更少。
如同薩瑞奇和威爾遜20年前所獲得的結果,許多人類學家這次也是群情嘩然,不肯相信肯恩和威爾遜的研究結果。當時對人類的進化有個普遍的看法:我們的祖先大約在200萬年前離開非洲,後來才在舊世界定居。根據這樣的理論模型,人類族譜的長度應該是他們所推算的13倍。肯恩和威爾遜的看法被媒體稱為「夏娃假說」(The Eve Hypothesis),或沒那麼誇張的「非洲起源說」(Out of Africa)。他們並沒有否定在更古老的時代所發生的遷移活動,而是暗示現代人抵達歐洲后,取代了早期的「類人」(hominid)族群,這些類人的祖先在將近200萬年前離開非洲來到了歐洲。200萬年前從非洲向外擴散的直立人(Homo erectus)在舊世界四處遷徙,並在70萬年前左右促成尼安德特人的興起,因此尼安德特人其實是這些直立人在歐洲的後代。後來,頂多在大約15萬年前,智人,或說現代人(他們也是直立人的後代,但是一直在非洲進化,沒有離開過)和久遠年代以前他們的直立人祖先一樣,選擇踏上了離開非洲的浩大旅程。我們在本章開頭已經看到,尼安德特人沒有和剛抵達歐洲的族群混種,這和現代人遇到直立人時的情形似乎相同。無論他們在什麼地方相遇,現代人都會取代直立人。最後一批尼安德特人在2.9萬年前左右消失,代表了最後一批非現代人的直立人後裔步入滅絕。
事實上,這種隨機滅絕的情況在統計學上是不可避免的。然而,這種情況的發生速率通常很慢,以至於惟有在很長的時間后才會變得明顯。不過有時瓶頸(人口規模銳減的時期)會大幅加速這個過程。在島上的族群歷史中,剛開始只有三對夫婦(六人),所以很可能在單單一代中便失去Smith和Watson這兩支,因為他們只生女兒或無法生育的幾率蠻高的。在大族群里,這種家系突然消失的情形不會發生;這在統計上是難以想像的事,如果一個族群里有許多Smith夫婦,很難想像他們全都只生育女兒或完全無法生育。惟有在經過許多世代后,家系逐漸縮小的效應才會慢慢浮現。事實上,我們假設的姓氏消失過程真的在南太平洋發生過:1789年著名的「叛艦喋血記」事件中,英國船艦「豐饒號」(Bounty)的6名叛變船員帶著他們的13位塔希提島新娘佔領了皮特肯島(Pitcairn Island)。在7個世代內,島上的姓氏就減少到了3個。
針對一個區域,詳細地研究該地Y染色體和線粒體DNA的變異,也可以看出在殖民過程中所造成的性關係與婚配習慣。比方說,在維京人抵達前,冰島原本無人居住,但在對線粒體DNA與Y染色體進行比較后,我們發現有顯著的不對稱情形。大多數的Y染色體都如預期一樣是來自斯堪的納維亞人(Norse,通稱北歐人),但大部分的線粒體DNA類型則是來自愛爾蘭。斯堪的納維亞人在殖民冰島時,顯然帶著愛爾蘭婦女。可惜我們無法從線粒體DNA的數據中得知當時愛爾蘭婦女的感受。
帕博的實驗室也率先釆取一項看來頗有希望的創新方法,尋找其他可能為重要差異編碼的基因。帕博利用可決定特定組織內有哪些基因被激活的DNA微陣列(參見第八章),比較人類、黑猩猩和獼猴體內三種組織——白血球、肝組織和腦組織——的基因表現模式(亦即哪些基因被啟動)。人類和黑猩猩的親緣關係近,結果發現人類與黑猩猩的白血球和肝組織在基因表現上也很接近,如同預期。但腦組織的基因表現卻截然不同:人類的腦和黑猩猩及獼猴的腦非常不同。或許這也沒什麼好驚訝的,大多數的人不需要有大堆儀器的實驗室,也知道人腦和黑猩猩的腦顯然不同。這個研究的意義在於提供在人腦與黑猩猩腦中表現不同的基因清單。但這也只是開端而已。就算找到基本機制的完整清單,我們也無法明確了解它們是「如何」使我們和黑猩猩變得不同。我們之所以為人,絕不是詳細列出受控的分子就能說明的。是在搜尋人之所以為人的遺傳基礎之際,至少現在我們已經可以開始編纂一份包含各種可能原因的清單。
克林斯在談到他第一眼看到定序結果時說:「那是一種無法形容的興奮感,好像有什麼東西爬上我的背脊。」雖然,如同原先的憂慮,有些序列證明已被污染,但他也在一些其他的序列上看到很不可思議的結果:這些序列跟現代人的序列不同,但又具有令人好奇的相似處。把片段拼湊起來后,他重建出尼安德特人的線粒體DNA,總共有379個鹼基對。但是當時賓州的結果還沒出來。不過最後證明他們的序列是相同的,都有一模一樣的379個鹼基對。克林斯回憶說:「直到那時,我們才開香檳慶祝。」
不論好壞,歷史向來是男性遷移的編年史,而非女性的。一般而言,男性出外是為了掠奪或建立帝國:亞歷山大大帝從馬其頓揮師入侵印度北部地區;維京人縱橫大海,從斯堪的納維亞到冰島,甚至更遠的北美洲;成吉思汗率領騎兵越過中亞的大草原等都是實例。但是即使沒有戰爭作為旅行的原因,我們仍會認為在人類社會中,男性的移動性比較大。傳統上男性負責狩獵,所以會到離家很遠的地方,而在傳統的釆集/狩獵社會,女性大多在家附近採集食物,養兒育女。西斯塔德因此認為,男性應該是傳播人類基因的原動力。研究結果的數據卻證明他錯得離譜:一般而言,女性的移動性比男性大八倍!
研究遺傳變異在各大洲的分佈情況,再加上考古信息,提供了我們一些人類祖先在全球擴張的細節。人類沿著亞洲邊緣,從如今的印度尼西亞群島、新幾內亞到澳洲的遷徙旅程,大約是在6萬年前完成的。要抵達澳洲,必須橫越數個浩大的水域,這顯示我們的祖先在這個早期階段已經開始使用船隻。現代人類大約是在4萬年前抵達了歐洲,又在大約1萬年後越過北亞,包括日本。
然而,尼安德特人引發的最大爭議,主要在於他們跟我們的親緣關係,而不在於他們有多聰明。我們是他們的後代嗎?古生物學顯示,現代人大約在尼安德特人消失時抵達歐洲。這兩個群體之間是否曾經通婚,還是尼安德特人被現代人消滅了?由於這些事件發生在遠古時代,現存證據又殘缺不全(只有一些零星的骨骸),這類的爭論還會延續許久,讓學院派的古生物學家和人類學家樂此不疲。在尼安德特人典型的厚骨和現代人較輕的骨頭之間,是否有另一種骨頭存在?這類骨頭有可能是這兩個群體的混種,一個「失落的環節」。但它們也可能完全屬於尼安德特人,只不過是某一位骨頭特別輕的尼安德特人,或者它們有可能完全屬於現代人,只不過是一位骨頭特別厚的現代人。
尼安德特人DNA的研究已經證明,我們和他們在遺傳上明顯不同。但就整體而言,有關人類進化的分子學研究似乎朝相反的方向發展:它們顯示我們和自然界其餘生物的遺傳關係近得驚人。事實上,分子資料經常對以往有關人類起源的假設提出質疑,甚至推翻它們。
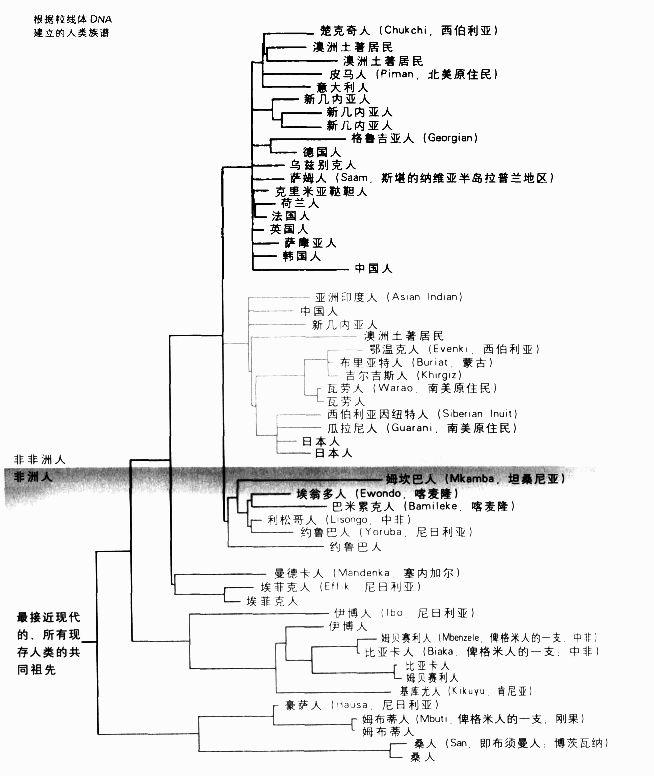
根據粒線體DNA建立的人類族譜
根據定義,史前時代是指有書寫紀錄以前的時期,但是我們卻發現,我們祖先的個別旅程早就記錄在了我們每個人的DNA序列中。新興的分子人類學運用不同族群的遺傳變異模式來重建人類的拓殖史,藉以窺知人類的「史前時代」。
線粒體DNA和Y染色體的資料也證實了另一項有趣的發現,那就是南非的桑人(San)在人類族譜中的位置。他們在人類族譜中的分支長度是最長的,也就是最古老的。這絕非顯示他們比我們其餘的人「原始」,每一個人類跟與我們親緣關係最近的大猿之間,進化差距和分子差距都一樣。如果我們沿著譜系追溯到黑猩猩和人類最後一個共同祖先,我的譜系會有大約500萬年之久,而桑人的也一樣。事實上,在這段漫長歲月的大多數時間里,我們這兩個譜系都是相同的;一直到15萬年前,桑人的譜系才跟其他人類的譜系分開。
帕博的研究生克林斯(Matthias Krings)負責執行這個計劃。起初他不太樂觀,但是在對遺骨的保存狀態進行評估之後,初步的分析結果讓他決定大胆做下去。克林斯的做法跟一般的預期不同,他不是在細胞核中搜尋DNA,而是把焦點放在微小的線粒體(mitochondria)上。線粒體|位於細胞核外,遍布細胞各處,製造細胞所需要的能量。每個線粒體包含一小股DNA,長約1.66萬個鹼基對。由於每個細胞里有500個到100
九-九-藏-書0個線粒體,而細胞核里只有兩套基因組,所以克林斯知道就這些腐敗的尼安德特人遺骨而言,在線粒體中找到完整序列的可能性,要比從細胞核中找到完整序列高得多。此外,由於線粒體DNA(簡稱mtDNA)長久以來是研究人類進化的主要對象之一,所以他有充分的現代人序列可供比較。雖然膚色在人類歷史和個人生命上向來是很強大的決定因素(而且大多是負面的),但我們對膚色背後的遺傳學,所知卻少得令人驚訝。然而這可能不是因為科學本身的限制,而多半是政治介入科學的結果。在受到政治正確性壓制的學術界,即使研究膚色這類特徵的分子基礎都是禁忌。目前我們在這方面的少許知識,都是來自以前對混血兒的研究,在這些研究中發現了數個促成色素沉著的基因。但是我們對其他物種的了解,以及所有哺乳動物之間基本生化過程的相似性都顯示,真正的全貌沒這麼簡單。例如,我們已經知道許多基因會影響老鼠的毛皮顏色,而且人類可能也有對等的基因。不過到目前為止,我們只找出了兩個與人類色素沉著有關的基因:一個在發生突變時會引起白化症,另外一個是與紅髮和膚色淡(經常有雀斑)有關的黑皮質素受體(melanocortin receptor)。黑皮質素受體基因在歐洲人和亞洲人當中都有變異,但在非洲人中沒有變異,這意味著,在非洲,自然選擇對這個基因的突變非常不利,亦即不會選擇讓紅髮、皮膚白皙的人產生。完全缺乏色素的白化症患者,如今偶爾會在非洲族群中出現(可能是通過新生突變[de novo mutation]),但是由於他們對陽光極度敏感,所以處於非常不利的地位。
另一個可能也是由自然選擇決定的形態性狀是體型。在以散發體熱為優先考慮的熱帶氣候區,進化出了兩種基本體型。一是高而痩的尼羅河流域體型(Nilotic form),以東非的馬薩伊人(Masai)為代表,這種體型是把表面積對體積的比例增加到最大,以便促進散熱。另一是侏儒體型(Pygmy form),也是細痩型,但很矮,矮小的體型比較適合需要耗費體力的採集/狩獵生活,使行動時耗費的能量減到最少。畢竟,何必拖著龐大的身軀去尋找食物?相對地,在高緯度地區,自然選擇對能留存體熱的體型有利,亦即表面積對體積的比例較低的體型。因此歐洲北部的尼安德特人身材粗壯,如今北方國家的居民一般也是這樣。今日不同族群在運動表現上的差異,大概也可以歸因於這些體型上的差異,例如在跳高方面,高大的尼羅河流域體型比矮壯的體型更適合,應該也沒什麼好奇怪的了。
後續的研究證實了肯恩和威爾遜的結論。新的研究結果大多來自卡瓦利-斯弗扎(Luigi Luca Cavalli-Sforza)在斯坦福大學的實驗室,他是用遺傳學方法來研究人類學問題的先驅。卡瓦利-斯弗扎出身義大利米蘭的名門望族,對顯微鏡很著迷。他相當早熟,1938年16歲時就進入帕維亞大學(University of Pavia)醫學院就讀。他曾特別提及,「那是一個相當幸運的選擇」。否則他的另一個選擇是加入墨索里尼的軍隊。1951年我第一次遇到他時,卡瓦利-斯弗扎還是積極有為的細菌遺傳學家。但是一位研究生閑聊間無心的一句話,讓他從細菌遺傳學轉而研究人類遺傳學。這位研究生曾接受成為神職人員的訓練,偶然提及天主教會保存了超過三個世紀的詳細婚姻記錄。卡瓦利-斯弗扎發現這些記錄可以提供豐富的研究資源,因而逐漸轉向研究人類遺傳學,他可能是極少數通過天主教教會找到職業專長的人類遺傳學家之一。
分子生物學最偉大的任務當然是回答有關人類,以及人類這個物種起源的問題。但是,除了人類全體的故事,每一個人都會渴望知道其本身的故事。DNA也可以提供更個人化的譜系記錄,讓我們更加了解我們各自的祖先。我的DNA分子內可以說是記錄著我的進化傳承歷史,那是一個可以從不同層面來看的故事。我可以把我的線粒體DNA序列放進肯恩和威爾遜的人類族譜,或者我也可以更仔細地了解自己家族的過去。我的Y染色體和線粒體DNA將述說不同的故事:一個說的是我父親那邊,一個說的是我母親那邊的故事。
研究進化的分子學方法取決於兩個變數的相互關係:兩個物種(或族群)在進化上開始分歧后的時間長短,以及他們之間的分子差異程度。這個「分子鍾」(molecular clock)的邏輯很簡單。為了方便說明,讓我們假設有兩對同卵雙胞胎,一對男的,一對女的,每對雙胞胎的基因完全相同。我們讓兩對雙胞胎彼此婚配,然後讓這兩對夫婦各在一個荒島上獨居。從遺傳學觀點來看,這兩個島上的族群起初無法區分。現在讓這兩對夫婦及其後代在各自的荒島上繁衍數百萬年,等這段期間過去后,其中一個族群應該已經有了另一個族群沒有的突變,反之亦然。由於突變的發生速率慢,再加上個體的基因組大,有許多位置都可能發生突變,所以這兩個族群發生完全相同的突變幾乎是不可能的事。因此,當我們為每對夫婦的後代定序DNA時,會發現這兩對夫婦原本一模一樣的基因組已經累積了許多差異,也就是說這兩個族群已經有了遺傳變異,而且互相分離的時間愈長,差異愈大。
在獲得線粒體DNA和Y染色體的家族數據后,我們人類這麼相似的原因就昭然若揭了:我們共同的祖先存在的年代離現在並不遠;就進化的標準而言,15萬年的時間眨眼即過,在這麼短的時間內,是沒有辦法經由突變產生大量豐富變異的。
化石證據顯示,新舊世界的猴子(地球上兩種最主要的猴群),大約在3000萬年前從共同祖先開始產生分歧,因此威爾遜和薩瑞奇把新舊世界的猴子之間的免疫「距離」設定為3000萬年。如此一來,人類和進化上關係最近的黑猩猩及大猩猩等大猿(great apes)之間的距離又是多少?1967年,兩人發表了他們的估計,認為人類和大猿大約是在500萬年前開始分道揚鑣。他們的說法引起一陣嘩然,因為古人類學家向來認為,人類和大猿早在2500萬年前左右就已分離。傳統理論堅持,人類和大猿間的差異,顯然是在遠遠超過500萬年以上的時間中進化而成的。許多人認為光憑這一點,就足以證明這個伯克萊團隊新式的遺傳學方法沒有價值;他們宣稱,遺傳學家還是專心研究果蠅吧,把有關人類的研究留給人類學家!但是威爾遜和薩瑞奇還是撐了下去,而且後續的研究也證明,他們所定出的人類與猩猩在進化上的分歧時間相當精確。
肯恩和威爾遜發現人類族譜有兩個主要的分支,一支僅包含非洲的一些不同族群,但另一支則包含一些非洲族群和所有的其他族群。這表示現代人起源於非洲,也就是說我們所有人的共同祖先住在非洲。但這幾乎稱不上是新觀念。達爾文在注意到親緣關係跟我們最近的黑猩猩和大猩猩都是非洲原產的物種后,就曾推論人類也是在非洲進化的。在肯恩和威爾遜所建立的人類族譜上,最驚人和爭議最大的一點,在於人類族譜最早可以追溯至多久遠的年代。在對突變于進化中的累積速率作出一些簡單的假設后,就可以計算人類族譜的年代,也就是一直往回追溯至我們的曾曾曾曾曾曾……祖母的年代。肯恩和威爾遜獲得的估計值是15萬年,換句話說,即使現存親緣關係最遠的人,也在15萬年前有共同的祖先。
人類和黑猩猩的染色體非常相似。但是黑猩猩有24對染色體,我們則是23對。近來我們發現,人類的第2號染色體是由黑猩猩的兩個染色體結合而成。人類和黑猩猩的第9號染色體(人類的比較大)以及第12號染色體(黑猩猩的比較大)也有一些差異,染色體中的數個倒位(inversion,基因的排列順序反轉)也有所不同。這些染色體上的差異是否具有重要意義,目前還很難講。
有一種解釋指出,向來主要仰賴乳製品的族群進化出了乳糖耐受性(lactose tolerance)。但這個性狀的模式仍無法令人完全信服這種解釋,比方說,在中亞以畜牧為生、人人都以乳酪為食的族群,仍有乳糖不耐症。儘管我本身所屬的族群通常可以耐受乳糖,但我本身卻有乳糖不耐症。如果自然選擇對特定族群的乳糖耐受性有利,為什麼同一族群里不是所有的人都可以耐受乳糖?到目前為止,支持這種標準解釋最有力的證據在於傳統上以畜牧為生的非洲族群中,確實具有乳糖耐受性。我們可能永遠無法完全了解這個性狀的適應範圍,但是研究芬蘭族群的分子生物學家最近已經找出造成這個性狀的突變。雖然這不是攸關生死的問題,但是現在只要做個簡單的遺傳試驗,就可以得知一個新生兒在長大後有什麼選擇:終生不吃冰淇淋,還是長期忍受胃痛?
在有了這些性別特異性(sex-specific)遺傳資料(女性的線粒體DNA,男性的Y染色體)之後,就可以比較男性與女性的歷史。卡瓦利-斯弗扎的研究生西斯塔德(Mark Seielstad)挑的題目是比較兩性的遷移模式。這個研究的邏輯很簡單:假設在南非的開普敦市,有一個Y染色體發生突變,則該突變傳播至開羅的速度,就可以作為男性遷移速率的指標。同樣地,在開普敦市出現的線粒體DNA突變傳播至開羅的速度,可以用來測量女性遷移的速率。
為了更加了解自然選擇對遺傳變異模式(以及人類族譜的整體年齡)的影響,讓我們先假設下列的情況:在15萬年前,有一個早期原始人類族群擁有過多的線粒體DNA序列,如同今日的人類,結果在這些序列中,有一個序列發生了有利的突變——自然選擇偏好的突變。接著這個突變發生的頻率會增加,直到許多世代后,這個種群的所有成員都有了這個突變為止。由於線粒體內不會發生DNA重組,線粒體DNA之間沒有交換,自然選擇過程會對有利突變首度出現的那個序列造成整體影響,這個種群的每一個成員最後都會擁有相同的線粒體DNA序列。因此等到自然選擇的工作完成後,每一個個體都擁有了這個有利突變,而這個種群的線粒體DNA也不再有遺傳變異。然而,隨著時間逐漸推移,突變會再度發生,變異也再度增加,但是這些新的線粒體DNA序列終究都是源自於最早發生突變的那個序列:這就是族譜的趨同點,亦即所有序列最近代的共同祖先。這也正是肯恩和威爾遜發現的模式,但是在這個例子中,趨同點頂多隻代表進化對線粒體DNA的微調(fine-tuning)。