02 雙螺旋:生命之所在
那時,盧里亞嘗試替我安排卡文迪什實驗室的研究職位。卡文迪什位於劍橋大學,是科學界最著名的實驗室,盧瑟福(Ernest Rutherford)就是在這裏首先描述出原子的結構。當時那是布喇格的研究領域,而我則被安排跟英國化學家肯德魯(John Kendrew)實習,他的興趣是找出肌紅素(myoglobin)這種蛋白質的三維結構。盧里亞建議我儘快前往卡文迪什實驗室,因為肯德魯那時在美國,佩魯茨會審核我的資格。肯德魯與佩魯茨早先曾一起建立醫學研究委員會(Medical Research Council, MRC)作為生物系統結構研究的單位。
儘管這種轉型現象似乎與當時的了解相悖,但格里菲斷的觀察結果起初並未在科學界激起太大的漣漪。部分原因在於格里菲斯非常注重隱私,厭惡大型聚會,鮮少參加科學會議,有一次還是在別人強迫下才發表演講。那次他被同事架到計程車上,護送至演講廳,然後以含糊單調的聲調發表演說,談的是他在微生物領域所作的艱澀研究,完全沒有提及細菌轉型。幸好,並非所有人都忽視格里菲斯的突破。
來年春天,我不再研究DNA,反而繼續作那些戰前的研究,用卡文迪什實驗室強大的新X光束,研究鉛筆狀的煙草花葉病毒(tobacco mosaic virus)。
以我為例,薛定諤觸動了我的心弦,因為我對生命的本質也很感興趣。當時仍有少數科學家認為,生命仰賴全能的上帝賦予生命力。不過如同我大多數的老師,我也鄙視生機論的觀念。如果這種「生命力」是大自然運作的主宰,我們勢必很難經由科學方法來了解生命。反之,一想到生命可能藉由一本以密碼寫成的指令書而永續長存,就令我神往不已。什麼樣的分子密碼能夠複雜到足以傳遞眾多的生命奇迹?又有什麼樣的分子秘密,讓染色體在複製時都能複製出一模一樣的密碼?

小而美:沃森和克里克在《自然》雜誌發表的短文,宣布他們發現了DNA的雙螺旋結構,篇幅只有一頁。同期也刊載了富蘭克林與威爾金斯較長的文章。
艾弗里對肺炎雙球菌糖衣般的莢膜也很感興趣。他複製格里菲斯的實驗,試圖分離出使R型菌變成S型菌的物質,找出它的特徵。1944年,艾弗里、麥克勞德與麥卡提公布了他們的研究結果:他們以精心設計的一組實驗明確地證實,DNA就是造成這祌變化的「轉化因子」(transforming factor)。艾弗里及其研究小組用試管培養細菌,而不是用老鼠,因此更容易在加熱致死的S菌細胞上,找出哪個化學物質是轉型因子。他們有系統地一一破壞經過加熱處理的S型菌的生化成分,看要摧毀哪一種成分,才能阻止轉型發生。

物理學家薛定諤。其著作《生命是什麼?》啟發了我投入基因領域。
在《生命是什麼?》中,薛定諤提議,生命的語言也許就像摩斯密碼,是一系列的點與線。這個講法倒是蠻接近事實的。DNA的語言是由A,T,G與C構成的線性序列。就像我們在謄寫書籍時,偶爾也會打錯字一樣,所有的A,T,G,C在沿染色體複製時,也會出現極少量的錯誤。這些錯誤就是遺傳學家近50年來所一直探討的突變。在英文中,將i變成a,Jim就會變成Jam,而在DNA中,將T變為C,ATG就變成了ACG。
這真是令人難忘的時刻,我們覺得這次肯定對了。這麼簡單美麗的構造,絕對錯不了。最令我們興奮的是鹼基序列沿著雙鏈排列的互補特性,只要知道一條鏈上的序列(鹼基的順序),自然就能推知另一條鏈上的序列。這顯然是細胞分裂前染色體在進行複製時,基因的遺傳訊息能精準重現的原因。

薛定諤認為,我們可以從儲存與傳遞生物信息的觀點來思索生命。因此,染色體只是信息的攜帶者。由於每個細胞都要容納這麼多的信息,因此這些信息必須壓縮成薛定諤所謂的「遺傳密碼腳本」(hereditary code-script),植入染色體的分子結構內。要了解生命,就必須辨識這些分子,破解它們的密碼。他甚至臆測,了解生命(包括找到基因)說不定能讓我們超越當時所知的物理定律。薛定諤這本著作的影響甚巨,日後許多在分子生物學這出大戲的序幕中成為要角的人物,包括我和克里克(他先前是物理學者)在內,都拜讀過這本書,而且深受感動。
此時我已經發現,DNA的密度測量證據比較傾向於雙鏈,而非三鏈的模型結構。因此,我決定尋找可能的雙螺旋體。身為生物學家,我偏好遺傳分子是由兩個,而非三個成分組成的概念。畢竟,染色體就像細胞一樣,數量是以複製成兩倍而非三倍的方式增加。
1954年夏天,這兩人在馬薩諸塞州伍茲霍爾(Woods Hole)的海洋生物實驗室結識,當時我在那裡講學。喝了不少馬丁尼之後,兩人決定他們應該搭擋研究科學。他們的合作結果贏得了「生物學上最完美的實驗」之稱。
同樣受到薛定諤那本著作感召的前物理學家本澤(Seymour Benzer)也出席了這場盛會。他立刻了解到我們的突破性發現,對於他的病毒突變研究具有重要意義。他發現現在他能夠以「摩根的孩子們」在40年前研究果蠅染色體的方式,來研究一小段的噬菌體DNA:他可以在基因圖上標出突變位置,就好像當年研究果蠅的先驅們在染色體上標出基因的位置。本澤跟摩根一樣,也得靠重組作用來產生新的遺傳組合。不過摩根可以利用現成的重組機制,即果蠅性細胞的產物。本澤則得用兩種不同的噬菌體同時感染一個的細菌宿主細胞,藉此引發重組機制。這兩種不同的噬菌體在重要區段有一個或多個突變的差異。在細菌細胞里,重組作用(分子片段的交換)有時會發生在不同的病毒DNA分子之間,產生新的突變置換,即所謂的重組體(recombinant)。在普渡大學(Purdue University)的實驗室里,本澤短短一年內就有驚人的豐富成果,他製作了噬菌體rⅡ基因的圖譜,顯示出一連串的突變(遺傳腳本上的所有錯誤)在病毒DNA上的線狀排列方式。這種「生命語言」既簡單又呈直線形狀,就像書頁上的一行文字。對於我在冷泉港所發表有關雙螺旋的演講,匈牙利物理學家齊拉特(Leo Szilard)的反應跟學術比較無關。他問我:「你能申請專利嗎?」有一陣子,齊拉特的主要收入來源是他和愛因斯坦共享的一項專利,後來他還試圖和費米(Enrico Fermi)一起申請1942年他們在芝加哥大學所建造的核子反應爐專利,但並未成功。不過當時和現在一樣,專利只授予有用的發明,當時沒有人想到DNA有什麼實際用途。於是,齊拉特建議,我們應該申請版權。
我立刻把論文帶到倫敦,告訴威爾金斯與富蘭克林,他們仍有成功機會。但深信D九_九_藏_書NA並非螺旋的富蘭克林甚至不想看這篇文章,以免受到泡令的螺旋觀念影響,連在我提出克里克的螺旋論點后也沒改變想法。倒是威爾金斯對我帶來的消息很感興趣;他現在更確定DNA是螺旋。為了證明這一點,他拿出一張6個月前,由富蘭克林的研究生葛斯林(Raymond Gosling)用X光拍下的照片,即所謂的B型DNA。在那之前,我甚至不知道有B型的存在。
一個月後在劍橋,佩魯茨向我保證,我很快就能精通必要的X光衍射理論,應該也很快就能融入他們人數不多的研究單位。我鬆了一口氣,因為他並沒有因為我的生物學背景而拒絕我。布喇格也沒有,他還從辦公室下來看了我一下。

DNA複製:雙螺旋先拉開,再各複製一股

克里克與卡文迪什實驗室的X光管
我一回到哥本哈根,立即拜讀了泡令有關α螺旋的研究。令人驚訝的是,他並非根據X光衍射的實驗數據推論出模型,而是根據身為結構化學家的豐富經驗,大胆推論哪種類型的螺旋結構最符合多肽鏈的化學特性。泡令製作蛋白質分子不同部分的比例模型,找出可能的三維結構。他將問題簡化成一種三維拼圖遊戲,既簡單又聰明。
原本我對這場研討會的期望不高,因為我認為要解開蛋白質或DNA的三維結構,起碼還要10年光景。從早期令人失望的X光照片看來,要通過X光來解開DNA的秘密,尤其不可能。會有這種結果也是很自然的,因為當時大家都預期,每個分子的DNA序列應該不盡相同。在表面構造不規則的情況下,DNA細長的分子鏈勢必不可能按照規律重複的模式整齊地排列,X光分析自然也無法成功。

DNA的化學骨幹
當我回到劍橋,向布拉格報告B型DNA的消息之後,他認為不應再禁止我和克里克研究DNA,而且很希望DNA的結構能由大西洋這一岸破解。於是我們再度著手建構模型,設法把已知的DNA基本成分湊成螺旋結構。這些基本成分就是分子骨幹以及4個不同的鹼基(腺嘌呤、胸腺嘧啶、鳥嘌呤與胞嘧啶)。我委託卡文迪什的工廠替我們做一套錫制的鹼基模型,但是他們的製造速度對我來說不夠快,最後我只得拿硬紙板來剪出粗略的模型。
威爾金斯在演講中展示一張最近拍到的X光衍射圖,上面有許多明確的反射影像,顯示它是高度規則的結晶體。由此可以推論出,DNA必定具有規則的結構,只要能解開此結構,就可以揭露基因的本質。我立即開始幻想自己搬到倫敦去,協助威爾金斯找出這個結構。演講過後,我去找他談話,但卻一無所獲。他只對我表示,未來還有更多艱辛的工作要做。
DNA不再只是一個籠統的縮寫名詞,我們必須徹底解開它的分子結構和所有的化學細節才行。
完成論文後,我發現自己別無選擇,只能到可以讓我研究DNA化學組成的實驗室。然而,不幸的是,由於我幾乎毫無理論化學的基礎,實在不夠格轉戰任何以有機化學或物理化學來進行艱難實驗的實驗室。後來在1950年秋,我拿到博士后研究獎學金,到生化學家開爾卡(Herman Kalckar)在哥本哈根的實驗室作研究。當時他正在研究構成DNA的小分子的合成作用,但我很快就發現,他的生化方法永遠無法解開基因的本質。在他的實驗室多待一天,就會晚一天了解DNA如何攜帶遺傳信息。
一直到1944年,DNA才成為遺傳界的焦點,當時艾弗里(Oswald Avery)在紐約洛克菲勒研究所(Rockefeller Institute)的實驗室發表報告說,肺炎病菌的外膜組成可以改變。這個結果出乎他和麥克勞德(Colin Macleod)與麥卡提(Maclyn McCarty)這兩位資歷較淺的同事意料。
富蘭克林不理會這張照片,把注意力集中在A型DNA上,因為她認為研究A型比較可能獲得有用的資料。B型DNA的X光圖是一個清晰的十字。既然克里克和其他人早已推論出,這類的反射圖案是由螺旋所造成,這項證據清楚說明DNA必定是螺旋狀!事實上,儘管富蘭克林持保留態度,但這個發現並不出人意表。幾何學本身就顯示,螺旋結構是一長串重複的單元(例如DNA的核苷酸)最合理的排列方式。不過我們仍不知道這個螺旋的外觀,也不知道它含有多少鏈。
解開雙螺旋:1953年6月沃森在冷泉港實驗室發表演說
同時,惟一兩位具有遠見、知道要尋找DNA三維結構的化學家,又犯了策略上的錯誤:富蘭克林不願建構模型,泡令則忽略了有關DNA的現有文獻,特別是查加夫所發表的DNA鹼基組成數據。1952年巴黎的生化會議后,泡令和查加夫還曾同船越過大西洋,兩人卻不投緣。泡令習慣於自己總是對的,而且相信自己可以用基本原理來解開任何化學問題。他的自信通常很有道理。冷戰期間,他是批評美國核武器發展計劃的主要人士,有一次在發表演說后,FBI警員質問他怎麼知道原子彈含有多少鈽?泡令傲然答道:「沒人告訴我,我自己想出來的。」在其後的幾個月,克里克和我(雖然我沒那麼熱衷)樂得把我們的模型拿給川流不息、充滿好奇的科學家們看。然而,劍橋的生化學家並未邀請我們到生化大樓發表正式演說。他們戲稱這是WC,拿我們的名字縮寫開雙關語的玩笑。我們沒做實驗就找到雙螺旋,令他們惱怒。
雙螺旋的發現敲響了生機論的喪鐘。認真的科學家,甚至有宗教信仰的科學家都已發現,要對生命有完整的了解,不需要尋找新的自然定律。生命不過就是物理與化學——儘管是極為精密複雜的物理與化學。接下來的工作是要找出,生命如何上演出隱藏在DNA中的密碼腳本。細胞的分子機器如何讀取DNA分子攜帶的訊息?下一章將介紹,極度複雜的讀取機制如何引導我們深入了解生命的形成。
這個實驗的工作量很少,我有許多時間遊走劍橋眾多的圖書館。我在動物學系看到查加夫的論文,他發現在DNA中,腺嘌呤和胸腺嘧啶的數量大致相同,而鳥嘌呤則與胞嘧啶的數量差不多。在聽到這一比一的比例后,克里克想到,在DNA複製時,腺嘌呤和胸腺嘧啶是否互相吸引,而胞嘧啶與鳥嘌呤之間是否也存有類似的吸引力。若是如此,DNA「親代」鏈上的鹼基序列(例如ATGC)應該會與「子代」鏈上的互補(亦即TACG)。
克里克對重要問題的錯綜複雜總是非常著迷。小時候,他老愛問問題,被問膩的雙親只好買一套兒童百科全書給他,希望能滿足他的好奇心,結果這反而讓他沒有安全感,他告訴母親,他怕長大時,所有的事物都已被人發現,而他將無事可九*九*藏*書做。母親向他保證,日後一定還會有一兩件事等著他發現。事後證明她說的很准。
因此,當我聽到來自倫敦國王學院(King's College)生物物理實驗室的英國人威爾金斯(Maurice Wilkins)在最後發表有關DNA的演講時,不禁驚喜交加。34歲的威爾金斯是物理學家,戰時曾參與制造原子彈的曼哈頓計劃(Manhattan Project)。對他和許多參与這個計劃的科學家而言,原子彈投到廣島和長崎,應該是他們研究工作的最高成就,結果這卻造成他們的理想破滅。威爾金斯曾考慮完全放棄科學,到巴黎去當畫家,但生物學引起了他的興趣——他也讀過薛定諤的書。當時他正設法用X光衍射法解開DNA的秘密。
1947年秋,我到印第安納大學,計劃以研究基因作為博士論文的題目,那時我們經常討論艾弗里的論文。當時已沒有人懷疑他實驗結果的正確性,而洛克菲勒研究所也有更多的研究結果出爐,顯示蛋白質不太可能是細菌轉型過程中的遺傳因子。至此,化學家終於把下一次重大突破的目標放在DNA上。英國劍橋精明幹練的化學家托德(Alexander Todd)迎接挑戰,開始鑒定連結DNA核苷酸的化學鍵。到了1951年初,他的實驗室證實這些連結總是相同的,亦即DNA分子的骨幹非常規則。同一時期,在奧地利出生,後來逃到美國,任職于哥倫比亞大學的內外科學院的查加夫(Erwin Chargaff)使用濾紙層析法(paper chromatography)這種新技術,萃取出多種脊椎動物與細菌的DNA樣本,測量DNA內四種鹼基的相對含量。他發現,有些物種的DNA以腺嘌呤與胸腺嘧啶居多,有些物種則是鳥嘌呤與胞嘧啶較多。因此,有可能任何兩個DNA分子的組成都不同。
泡令跟査加夫一樣,也參加了國際生化會議,那時的大新聞是噬菌體研究小組的最新結果。冷泉港的赫爾希與蔡斯(Martha Chase)才剛證實了艾弗里的轉化因子:DNA就是遺傳物質!赫爾希和蔡斯證明,進入細菌細胞的只有噬菌體病毒的DNA,它的蛋白質鞘(protein coat)留在外面。看來如果我們想揭開基因的本質,勢必得了解DNA的分子。在赫爾希與蔡斯成為大家的話題之後,我確定泡令也會將他的才華與化學知識,全力投注在解決DNA的問題上。
噬菌體小組認為,如同所有的病毒,噬菌體其實就是赤|裸裸的基因(病毒就只是一個蛋白質外鞘包著核酸),並依據這個想法來規劃研究計劃。這個概念是想像力豐富的美國遺傳學家穆勒(Herman J. Muller)於1922年首先提出的,三年後,他證實X光會引起突變。但是直到1946年,穆勒來印第安納大學任教不久之後,才獲得了遲來的諾貝爾獎。事實上,正是因為他在印大,我才會到這裏就讀。穆勒是在摩根的手下展開他的職業生涯的,遺傳學在20世紀前半葉的發展過程,沒人比他更清楚,我在第一個學期時就對他的講學非常著迷。然而,他在果蠅方面的研究,對我而言似乎屬於過去,而非未來,因此我只短暫考慮過請他指導我的論文。後來我選擇盧里亞的噬菌體實驗,這個實驗做起來比果蠅快得多:噬菌體的遺傳雜交(genetic cross)子代在隔天就可以進行分析。
我和克里克相遇的那個早晨,他聽說我來劍橋是為了大量學習有關晶體學的知識以便破解DNA的結構,大為高興。不久之後,我就請克里克談談他對使用泡令的模型建構法來破解DNA結構的看法。我們是不是還得做許多年的衍射實驗,才能實際下手去建構模型?為了加快我們研究DNA結構的速度,克里克邀請自戰後就認識的朋友威爾金斯,在星期天從倫敦過來共進午餐,這樣我們就能得知自從那不勒斯的演講后,威爾金斯還有哪些進展。

讓一切運作的原理:鹼基由氫鍵連結的互補配對
就在我連續碰壁時,加州理工學院享譽國際的化學家泡令(Linus Pauling)宣布獲得了重大的成就:他發現蛋白質里氨基酸鏈(稱為多肽[polypeptide])的排列結構,並且將這個結構取名為α螺旋(alpha helix)。這個突破會由泡令獲得其實並不意外,他是科學界的超級巨星。他所著的《化學鍵的本質》(The Nature of the Chemical Bond)奠立了現代化學的基礎,被當時的化學家奉為聖經。泡令非常早熟,他在俄勒岡州長大,父親是位藥劑師。他9歲時,父親曾寫信給《俄勒岡人》報,希望對方能提供他那好學不倦的孩子可以閱讀的書籍,還說他兒子已經讀完聖經和達爾文的《物種起源》。泡令的父親不幸早逝,家中經濟陷入困境,但這位前途無量的年輕人仍然完成了學業,相當難能可貴。
早在1953年,泡令就發表過描述DNA結構的論文。我急切地拜讀了大作,發現他提出的是三鏈模型,以磷酸糖的骨幹形成稠密的中央核心。乍看之下,它跟我們在15個月前所做的拙劣模型類似。但是,泡令沒有採用帶正電的原子(例如Mg2)來穩定帶負電的骨幹,而是採取非正統的做法,以氫鏈來連接磷酸鹽。不過,看在身為生物學家的我眼中,這種氫鍵所需的極酸狀態從不曾見於細胞內。我發瘋似的衝到托德在附近的有機化學實驗室,立刻就證實了我的看法:不可能的事居然發生了!全世界最優秀或至少最著名的化學家竟然弄錯了化學基本原理。實際上,泡令等於把DNA里代表酸的縮寫A除掉了。我們研究的對象是脫氧核糖核酸,但是他所提出的結構甚至不屬於酸類。
在薛定諤于都柏林演講的年代,大多數的生物學家都認為,最終科學界會證明蛋白質是遺傳指令的主要攜帶者。蛋白質是由20種不同的建構單元(氨基酸)所組成的分子鏈。由於氨基酸沿分子鏈排列的順序可以說有無限多種,因此原則上蛋白質是有可能隱含造成生命如此多樣的密碼信息的。雖然DNA就位於染色體上,為世人所知也有75年之久,但當時並未將DNA視為密碼腳本的可能攜帶者。1869年,在德國工作的瑞士生化學家米舍(Friedrich Miescher),從當地醫院沾滿膿的繃帶上分離出一種物質,並稱之為「核素」(nuclein),膿大多由具有細胞核的白血球構成(紅血球沒有細胞核),因此也具有包含DNA的染色體,米契爾等於在無意間發現了DNA的良好來源。稍後當他發現惟有在染色體里才找得到「核素」時,就知道自己有了重大發現。1893年,他寫道:「遺傳確保形態能世代延續,而這一切就隱藏在比化學分子還深的層次。它隱藏在結構化的原子群組內。因此,我支持化學遺傳論。」
我們一回到劍橋,我就安排卡文迪什的機械部門建造磷的原子模型,以便用於建造DNA里磷酸糖骨幹的片段。等這些模型做好后,我們開始測試骨幹在DNA分子中央彼此纏繞的不同方法。它們規則重複的原子結構,應該會讓原子形成一致且重複的構造。我們聽從威爾金斯的直覺,把重點放在三鏈模型上。當其中一個模型看似很有可能是答案時,克里克打電話給威爾金斯,宣稱我們可能找到了DNA的模型。
格里菲斯從參与公共衛生的經驗中得知,https://read.99csw•com單一病人身上有時能分離出多種類型的菌株,因此他很好奇,不同類型的菌株在實驗鼠身上會如何交互作用。後來他在一種組合上有了重大發現:當他將加熱殺死的S型菌株(已變得無害)及正常的R型菌株(原本就無害)同時注入老鼠體內時,老鼠會死亡。兩種無害的菌株在混合后,怎麼可能變得致命呢?後來他分離出死老鼠身上的肺炎雙球菌,發現裏面有活的S型菌,線索於是出現。無害的R型菌似乎會從已死的S型菌取得不明物質;無論此物質為何,它顯然會使R型菌在有加熱致死的S型菌存在時,轉型為活的殺手型S菌。格里菲斯從死老鼠身上培育出數代的S型菌,證實這種變化的確存在:這些細菌繁殖成S型菌,如同任何正常的S型菌株。注入老鼠體內的R型菌真的發生了「遺傳」變化。
不過,我在哥本哈根的那一年仍然獲益良多。為了避開丹麥寒冷的春天,我在四五月間前往義大利那不勒斯動物研究所。在那裡的最後一周,我參加了一場小型研討會,主題是以X光衍射法(X-ray diffraction)決定分子的三維結構。X光衍射法可以研究任何能夠形成晶體的分子的原子結構。X光在轟擊晶體后,會在撞到原子時彈開而散射。從X光的散射圖形可以獲得有關分子結構的信息。但是只靠X光,尚不足以解決結構的問題,還需要所謂「相分配」(phase assignment)的額外信息,來處理分子的波性質(wave properties)。要解決「相」的問題並不容易,當時只有膽量最大的科學家願意麵對這種挑戰。以衍射法成功研究的對象,大多是比較簡單的分子。
我們在4月初將發現雙螺旋的報告交給《自然》(Nature)雜誌,並於3個星期後,也就是1953年4月25日刊出。同期還有兩篇由富蘭克林與威爾金斯執筆、篇幅較長的論文,兩篇文章都認為我們的模型大致正確。6月,我在冷泉港實驗室的病毒研討會上,首次就我們的模型提出報告。德爾布呂克親自出馬確保了我在最後一刻受邀發表演說。我帶了在卡文迪什製造的三維模型,參加這場精英薈萃的會議,我使用的腺嘌呤-胸腺嘧啶鹼基對模型是紅色,而鳥嘌呤-胞嘧啶鹼基對則是綠色。
威爾金斯表示,他認為DNA的結構是螺旋狀,由數條鏈接的核苷酸互相纏繞而成,而惟一尚待解決的問題在於鏈的數目。當時威爾金斯根據他測量的DNA纖維密度,認為應該有三條核苷酸鏈。他迫切地想著手建造模型,卻碰上了一個障礙:剛加入國王學院生物物理學研究單位的富蘭克林(Rosalind Franklin)。
DNA分子會像拉鏈一樣「拉開」,形成獨立的兩股。每一股都可以作為新股合成時的模板,於是一條雙螺旋就變成了兩條。

梅索森站在超離心機旁,這是「生物學上最完美的實驗」所使用的重要設備。
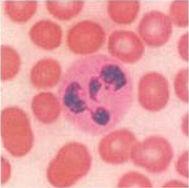
搜尋遺傳物質:左圖為顯微鏡下用DNA著色劑處理過的血球細胞。為使攜氧能力達到最大,紅血球沒有細胞核,因此也沒有DNA。但是在血流里搜尋入侵者的白血球有細胞核,內含染色體。
然而,在雙螺旋的拼圖中,還是少了一塊:對於DNA「像拉鏈一樣拉開」的複製方式,我們還需要以實驗來證實。德爾布呂克就不相信我們的概念,他喜歡雙螺旋的模型,但是擔心把雙螺旋拉開,會產生可怕的打結情況。5年後,泡令以前的學生梅索森(Matt Meselson)及同樣聰明的研究噬菌體的年輕學者史塔爾(Frank Stahl),發表了一個簡單明了的實驗結果,一舉掃除了這類疑慮。
直到1962年,克里克、威爾金斯和我才獲得諾貝爾生理醫學獎。在此4年前,富蘭克林已經因卵巢癌而不幸早逝,享年37歲。那時克里克已和她成為親密的同事與真正的好友。富蘭克林動了兩次手術,但都未能遏阻癌細胞蔓延,之後她曾回到劍橋,在克里克和他太太歐蒂莉(Odile)的照顧下療養。

布拉格(左)與拿著α螺旋模型的泡令合影
α螺旋相當美麗,但現在的問題在於它是否正確。短短一星期後,我得到了答案。發明X光晶體學(X-ray chrystailography)的1915年諾貝爾物理學獎得主布喇格爵士(Sir Lawrence Bragg)來到哥本哈根,興奮地宣布,比他資淺的同事、奧地利籍化學家佩魯茨(Max Perutz)巧妙地用合成多肽證實了泡令的α螺旋是正確的。對布拉格的卡文迪什實驗室而言,這是個苦樂參半的勝利,因為前一年他們在論文中列舉多肽鏈可能具有的螺旋形態時,完全不得要領。
我在芝加哥大學三年級時,迷上了基因。我原本想當博物學家,向往日后能離開自幼成長的芝加哥南區,到沒有都市塵囂的地方發展職業生涯。讓我改變心意的,並不是某位難忘的老師,而是1944年出版的一本薄薄的小書《生命是什麼?》(What Is Life?),作者是奧地利籍的波動力學之父薛定諤(Erwin Schrodinger)。這本書輯錄了他前一年在都柏林的高等研究院(Institute for Ad-vanced Study)發表的數場演講。這麼偉大的物理學家竟會花時間寫生物學的書,引起了我的興趣。當時我和大多數的人都認為,化學與物理學才是「真正的」科學,而理論物理學更是科學翹楚。
第二天,威爾金斯與富蘭克林一起來訪,査看我們的成果。在這意外出現的競爭威脅下,他們倆為了共同目標而難得地暫時合作。富蘭克林立即挑出我們在基本概念上的錯誤。我記得她在報告時指出:DNA晶體幾乎不含水。其實是我自己弄錯了。由於才剛開始學晶體學,我把晶胞(unit cell)與不對稱單位(asymmetric unit)這兩個術語搞混了。其實,DNA晶體富含水分。因此,富蘭克林指出,光是要容納她在晶體內觀察到的水分子,骨幹就得在分子外面,而不是像我們所做的在分子中央。
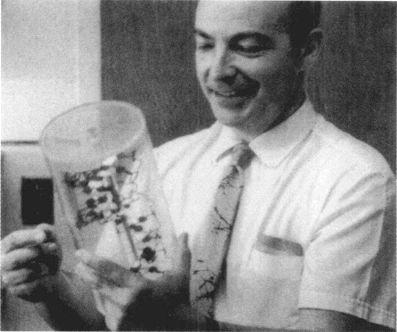
科恩伯格贏得諾貝爾獎,他手中拿著雙螺旋模型的複製品。
我知道先前將骨幹置於裏面,而鹼基懸挂在外的模型是錯誤的。諾丁漢大學提出的化學證據顯示,鹼基必須由氫鍵彼此連結,但這項證據一直被我忽略。如果鹼基位於分子中央的話,就只能按照X光衍射數據所顯示的規律方式形成這種鍵。但是它們怎麼會成雙作對呢?在錯誤的核酸化學教科書誤導下,我連續兩個星期毫無進展。幸好2月27日加州理工學院的理論化學家多納休(Jerry Donahue)到卡文迪什訪問,他指出教科書的錯誤。於是我改變了氫原子在硬紙板分子https://read.99csw.com模型上的位置。
富蘭克林才從法國回來,她在巴黎用X光晶體學的技術研究了4年的石墨。她接受聘任,加入國王學院的DNA計劃時,威爾金斯剛好不在。不幸的是,後來證明這兩人根本合不來。富蘭克林的個性率直,重視數據,威爾金斯則拘謹而勇於猜想,他們註定無法合作。在威爾金斯接受我們的午餐邀約前不久,他們兩人才大吵一次,富蘭克林堅持在她搜集更多衍射數據前,不能著手建立模型。他們倆顯然無法溝通,而在富蘭克林於11月初舉辦實驗室研討會之前,威爾金斯無從得知她的進展。不過若我們想參加這次研討會,威爾金斯很樂意邀我和克里克前往。後來克里克因故未能參加研討會,由我獨自前往,稍後再把我認為與DNA晶體有關的重要訊息告訴他。我特別根據記憶,描述了富蘭克林關於晶體重複與含水量的測量值。克里克聽了之後,開始在紙上繪製螺旋網格,並說就連我這種先前以賞鳥為業的人也可以應用他和科克蘭(Bill Cochran)及凡德(Vladimir Vand)提出的新螺旋X光理論,準確預測我們即將建造的分子模型有哪些衍射圖。
艾弗里的研究小組花了10餘年的時間,持續追蹤英國衛生部科學家格里菲斯(Fred Griffith)在1928年視察到的奇特現象。格里菲斯對肺炎很感興趣,潛心研究肺炎的致病菌——肺炎雙球菌(Pneumococcus)。當時已知肺炎雙球菌有兩種形態,依照它們在顯微鏡下的外觀而分為「平滑」(smooth)的S型與「粗糙」(rough)的R型。兩者不僅外觀不同,毒性也不同。將S型注入老鼠體內,幾天內老鼠就會死去,但是注入R型的老鼠則依舊健康。後來發現S型的細胞有莢膜,可以防止老鼠的免疫系統認出它是入侵者。R型的細胞沒有莢膜,因此會受到老鼠免疫系統的攻擊。
隔天早晨,1953年2月28日,DNA模型的重要特徵全都各就各位。它的兩條鏈由腺嘌呤-胸腺嘧啶,以及鳥嘌呤-胞嘧啶這兩對鹼基對之間的強氫鍵連在一起。克里克一年前根據查加夫的研究所得到的推論,真的是正確的。
11月的那一天實在不幸,讓未來蒙上濃濃陰影。富蘭克林更加堅定地反對建造模型,她打算繼續做實驗,不玩看似小孩玩具的原子模型。更慘的是,布拉格也說話了,叫我和克里克不要再嘗試建造DNA模型,後來還進一步決定,DNA研究應交由國王學院實驗室來做,劍橋只需繼續研究蛋白質即可。兩家同樣由醫學研究委員會贊助的實驗室居然彼此競爭,實在沒有道理。在無計可施下,我和克里克不情願地暫時罷手。
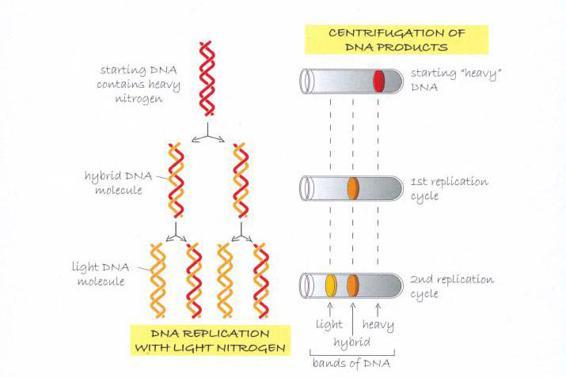
如果DNA的複製如同我和克里克的預測,是將雙螺旋拉開,然後各複製一股,那麼實驗所製造出的兩個「子代」DNA分子將會是混種,每個分子都包括一個重氮股(來自「親代」分子的模板股),以及一個輕氮股(用培養基製造出來的新股)。梅索森與史塔爾的離心處理程序完全證實了這種預測。他們發現離心試管分離出三個明顯的區段,分別是重—重、輕—輕,以及介於兩者之間、先重后輕的樣本。DNA的複製方式跟我們的模型所預測的完全相符。
梅索森和史塔爾用細菌所做的實驗
艾弗里錯失的,不僅是反擊同事、為自己的研究成果辯護的機會,他也錯失了獲得諾貝爾獎的機會,身為DNA是轉化因子的發現者,他其實很有獲獎資格。由於諾貝爾獎委員會在各個獎項頒發50年後會公布記錄,現在我們已經知道當時阻擋艾弗里獲得候選資格的人,是瑞典籍的物理化學家哈馬斯滕(Einar Hammarsten)。雖然哈馬斯滕的聲望主要奠基於他能萃取出質量絕佳的DNA樣本,但他仍然相信基因是某類尚未辨識出來的蛋白質。事實上,即使在發現雙螺旋后,一直到DNA轉型的機制完全公布前,哈馬斯滕仍堅持艾弗里不應獲得諾貝爾獎。艾弗里於1955年過世,若是他再多活幾年,肯定可以拿到諾貝爾獎。
31歲的富蘭克林是出身劍橋的物理化學家,她是個對自己的專業執著得不得了的科學家,在29歲生日時,她只要求訂閱自己所屬領域的技術期刊《晶體學報》(Acta Crystallogrsphica)作為生日禮物。她講求邏輯和精確,對於欠缺這些特質的人沒有什麼耐性。她不習慣快速發表措辭強烈的看法。一度將她的博士論文指導教授,未來的諾貝爾獎得主諾里什(Ronald Norrish)描述為「愚蠢、固執、奸詐、態度惡劣、專制」。在實驗室外,她是果決勇敢的登山家,來自倫敦的上流社會,相較於大多數的科學家,她屬於高尚的社交界。在工作台辛苦一整天後,她偶爾會脫下實驗室的外套,換上優雅的晚禮服,消失在夜色中。
為了完成博士論文研究,我在盧里亞的要求下跟隨他的步履,研究X光如何殺死噬菌體粒子。剛開始時,我希望能證明病毒死亡是因為噬菌體的DNA遭到破壞。但最後我不得不承認,我的實驗方法在化學上永遠無法獲得確切的答案,只能得到生物學上的結論。雖然噬菌體的確是裸|露的基因,但我知道噬菌體研究小組所要的深奧答案,惟有通過高深的化學才找得到。
現在終於到了我們繼續建構DNA螺旋模型的時候。泡令肯定不久就會發現,他的精心傑作出現了謬誤。我敦促威爾金斯不要再浪費時間,但是他想等到富蘭克林在該年春天稍後到另一家實驗室工作后才開始。她選擇離去,避開在國王學院的不愉快。在離開前,她奉命停止對DNA作更進一步的研究,並將許多衍射照片交給威爾金斯。
諾貝爾委員會向來不曾將單一獎項頒給超過三個人,倘若當時富蘭克林還在人世,他們勢必得面對要將這個獎頒給她或威爾金斯的問題。瑞典人可能會授予他們諾貝爾化學獎以解決這個難題。最後,化學獎頒給了佩魯茨和肯德魯,他們分別發現了血紅素與肌紅素的三維結構。
克里克很擅長說話,無論是哪種聚會,他總是眾人注意的焦點。在卡文迪什實驗室的走廊上,總是可以聽到他爽朗的笑聲。他是醫學研究委員會研究單位的專任理論家,每個月至少會提出一個新構想,而且只要有人願意聽,他總是很樂意花許多時間仔細解釋。
我在10月初抵達劍橋醫學研究委員會的研究單位,那年我23歲。我和35歲的前物理學家克里克共用生化研究室,他在戰時曾替英國海軍研究磁性水雷。戰爭結束后,克里克原本計劃留在軍方的研究機構,但在拜讀了薛定諤的《生命是什麼?》后,決定朝生物學發展。當時他在卡文迪什實驗室以研究蛋白質的三維結構為博士論文。

鹼基與骨幹就位,形成雙螺旋:(A)將DNA的兩股連結在一起的鹼基配對系統;(B)將分子的原子細節按比例呈現的「空間填充」模型。
氮原子(N)是DNA的成分之一,而且原本就有兩種不同的形態,一種較輕,另一種較重,因此梅索森和史塔爾能藉由標註DNA片段,追蹤DNA在細菌里的複製過程。起初所有的細菌都在含有重氮的培養基里培養,讓重氮進入DNA的雙股上。然後他們從這個培養菌中取出樣本,轉移至僅含輕氮的九*九*藏*書培養基,確保下一次DNA在複製時,只會用到輕氮。

富蘭克林在假期時熱愛登山運動,攝於歐洲阿爾卑斯山。

威爾金斯,攝於倫敦國王學院的實驗室
在印第安納大學,我加入一小群以物理學家和化學家為主、具有遠見的科學家之中,共同研究感染細菌的病毒「噬菌體」(bacteriophage,簡稱phage)的繁殖過程。後來我的博士論文指導老師,也就是在義大利受訓的醫師盧里亞(Salvador Luria),以及他在德國出生的理論物理學家好友德爾布呂克(MaxDelbrück),和美國理化學家赫爾希(Alfred Hershey)攜手合作,成立了噬菌體研究小組(the Phage Group)。在第二次世界大戰期間,儘管猶太裔的盧里亞是被迫離開法國投奔紐約,德爾布呂克因為反對納粹主義而逃離德國,但他們仍被視為敵僑,沒有資格和美國科學界共同為戰爭效力。雖然遭到排擠,但他們還是在各自的大學實驗室里努力工作(盧里亞在印第安納大學,德爾布呂克則在范德比爾特大學),並且接連幾年夏季都到冷泉港實驗室合作進行噬菌體實驗。1943年,他們與才華洋溢但沉默寡言的赫爾希合作,當時赫爾希正在聖路易的華盛頓大學研究噬菌體。
腺嘌呤的確與胸腺嘧啶連結,而鳥嘌呤也與胞嘧啶連結,但是它們並非通過平坦的表面形成分子三明治。當克里克抵達時,他很快就了解了狀況,並且認同我的鹼基配對(base-pairing)方式。而且他當下就發現,這會造成雙螺旋的雙股以相反方向連接。
這些原本都只是空想,直到1952年夏天,査加夫在前往巴黎參加國際生化會議時途經劍橋為止。我和克里克認為不需要了解4種鹼基的化學結構,但査加夫對這看法頗不贊同。又聽到我們說,如有必要的話,可以到教科書里查它們的結構時,他更是不悅。我只希望能證實查加夫的數據與DNA結構並不相關。不過克里克卻興緻勃勃地要做一些實驗,尋找腺嘌呤與胸腺嘧啶(或鳥嘌呤與胞嘧啶)在溶液中混合時,可能會形成的分子「三明治」。但是,他的實驗沒獲得任何結果。
大約同一時間,酶學專家科恩伯格(Arthur Kornberg)位於聖路易市華盛頓大學的實驗室也在分析DNA複製過程的生化細節。科恩伯格發展出一種新的合成DNA的無細胞系統,從而發現了一種稱做DNA聚合肽(polymerase)的酶,這種酶連接構成DNA的不同小單元,形成DNA骨幹的化學鏈。科恩伯格發現DNA酶合成機制,是驚人的重要大事,因此在這些實驗完成後不到兩年,他就於1959年獲得了諾貝爾生理醫學獎的殊榮。在公布得獎后,科恩伯格拿著我在1953年帶到冷泉港實驗室的雙螺旋模型複製品攝影留念。
無論從化學或生物學的觀點來看,雙螺旋都很合理。我們現在無須擔憂薛定諤所說,要了解遺傳密碼如何複製有可能需要新的物理定律,事實上,基因的組成與其他的化學作用並沒有兩樣。那天稍後,在緊鄰卡文迪什實驗室的鷹吧吃午餐時,向來愛說話的克里克忍不住告訴大家我們剛發現了「生命的奧秘」。我雖同樣激動,但寧可等到做出漂亮的三維模型時才炫耀。
在此時退出DNA研究,實在不是時候。泡令已寫信給威爾金斯,請他提供一份DNA晶體的衍射圖。雖然威爾金斯拒絕,表示自己需要更多的時間來解讀,但泡令其實也不見得要依賴國王學院的資料。如果他願意的話,大可在加州理工學院自行研究X光衍射。
首先他們使S型菌糖衣般的莢膜水解,但轉型仍舊發生,這證明莢膜並非轉化因子。接著他們使用兩種酶(胰蛋白酶[trypsin]與胰凝乳蛋白酶[chymotrypsin])的混合製劑,這兩種酶都會破壞蛋白質,結果S型菌的蛋白質幾乎全遭破壞。出乎他們意料,轉化仍然繼續發生。他們又嘗試一種會分解核糖核酸(ribonucleic acid, RNA)的核糖核酸酶(ribonuclease, RNase),但是轉化再次發生。RNA也是一種核酸,與DNA類似,而且可能與蛋白質的合成有關。最後他們把目標鎖定DNA,讓從S型菌取出的萃取物接觸會破壞DNA的脫氧核糖核酸酶(deoxyribonuclease, DNase),這次他們總算命中目標,R型菌不再轉型為S型菌,轉型因子就是DNA。
然而,數十年後,化學仍無法分析龐大複雜的DNA分子。一直到20世紀30年代,科學家才證明DNA是由四種不同的化學鹼基所構成的長分子,即腺嘌呤(A)、鳥嘌呤(G)、胸腺嘧啶(T)與胞嘧啶(C)。不過在薛定諤發表演說的年代,科學界尚不明白DNA分子上的這些次單位(稱為脫氧核糖核苷酸[deoxynucleotide])在化學上如何鏈接,也不知道DNA分子的四種化學鹼基序列是否有差異。如果DNA真的是薛定諤所謂的密碼腳本,那麼這種分子應該有極多種不同的形式。不過在當時,一般仍認為整條DNA鏈有可能是由一個簡單序列(例如AGTC)一再重複出現而構成的。
化學家托德是最早看到我們這個模型的人之一。基因的本質如此簡單,讓他非常驚喜。然而,稍後他必然曾捫心自問,為什麼自己的實驗室在建立DNA鏈的一般化學結構后,未能進一步研究這些鏈在三維空間的組成方式,反而讓由生物學家和物理學家組成的雙人組找出這種分子的本質,這兩人對大學程度的化學甚至都不是那麼了解。然而話說回來,這正是我們成功的關鍵,至少是部分關鍵:我和克里克之所以能率先獲得雙螺旋的結論,正是因為當時大多數的化學家認為,DNA的分子太大,無法用化學分析來了解。

A型DNA與B型DNA的X光照片,分別來自威爾金斯與富蘭克林。分子結構的差異是由每個DNA分子水含量的差異所造成。
他們使用離心技術,按照重量的些微差異來分離分子。在離心旋轉后,較重的分子會比較輕的分子落到更接近試管底部之處。
艾弗里、麥克勞德與麥卡提在1944年2月提交研究報告后,科學界反應不一,部分原因在於這項發現太過驚人。有許多遺傳學家接受他們的結論,畢竟DNA在每個染色體上都找得到,它為什麼不能是遺傳物質?但是,也有許多生化學家對DNA分子是否複雜到能儲存龐大的生物信息,表示存疑。他們仍舊認為,最終會證實同為染色體構成要素的蛋白質才是遺傳物質。其實也難怪生化學家會這麼想,因為基本上蛋白質有20個氨基酸字母可以編碼龐大的信息,這要比只有4個核酸字母的DNA容易得多。跟艾弗里同在洛克菲勒研究所任職的蛋白質化學家墨斯基(Alfred Mirsky)更是激烈反對DNA是遺傳物質的說法,不過那時艾弗里已不再活躍于科學界,洛克菲勒研究所強制他在65歲時退休。